Article Text

Abstract
Objective The colonic microbiota is altered in patients with colorectal cancer (CRC). We investigated the microbiota composition of patients with colon cancer compared with controls devoid of neoplastic or inflammatory disease and the potential to modify the colonic microbiota with probiotics.
Design Biopsy samples were obtained from the normal mucosa and tumour during colonoscopy from 15 patients with colon cancer. Subsequent patient-matched samples were taken at surgery from the tumour and nearby mucosa from the patients with cancer, eight of whom had received two daily tablets totalling 1.4×1010 CFUs Bifidobacterium lactis Bl-04 and 7×109 CFUs Lactobacillus acidophilus NCFM. Faecal samples were obtained after colonoscopy prior to starting the intervention and at surgery. In addition, 21 mucosal biopsies from non-cancer controls were obtained during colonoscopy followed by later faecal samples. The colonic and faecal microbiota was assessed by 16S rRNA gene amplicon sequencing.
Results The tumour microbiota was characterised by increased microbial diversity and enrichment of several taxa including Fusobacterium, Selenomonas and Peptostreptococcus compared with the control microbiota. Patients with colon cancer that received probiotics had an increased abundance of butyrate-producing bacteria, especially Faecalibacterium and Clostridiales spp in the tumour, non-tumour mucosa and faecal microbiota. CRC-associated genera such as Fusobacterium and Peptostreptococcus tended to be reduced in the faecal microbiota of patients that received probiotics.
Conclusions Patients with colon cancer harbour a distinct microbiota signature in the tumour tissue and nearby mucosa, which was altered with probiotic intervention. Our results show promise for potential therapeutic benefits in CRC by manipulation of the microbiota.
Trial registration number NCT03072641; Results.
- COLORECTAL CANCER
- PROBIOTICS
- INTESTINAL MICROBIOLOGY
- TUMOUR MARKERS
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Summary box
What is already known about this subject?
▸ The risk of colorectal cancer (CRC) is strongly associated with lifestyle factors, including a dietary component which may be mediated by the intestinal microbiota.
▸ The intestinal microbiome is altered in patients with CRC, and there is a strong interest in identifying potential microbial markers for CRC.
▸ Specific probiotic bacteria have been shown to modulate inflammation and reduce tumour proliferation in animal models of carcinogenesis and may offer therapeutic benefits for CRC patients.
What are the new findings?
▸ The colon cancer-associated microbiota exhibits a distinct signature characterised by increased mucosal microbial diversity and differential abundance of specific bacterial taxa compared with non-cancer controls. Oral-associated pathogens are over-represented in colon cancer tumours and tend to co-occur.
▸ Although more difficult to obtain, intestinal mucosa samples, rather than faecal, provide a more comprehensive assessment of microbiota changes in colon cancer. Peptostreptococcus was over-represented in both mucosal and faecal samples and shows promise as a CRC marker.
▸ The colon cancer-associated microbial signature was modified by probiotic intervention and was characterised by the enrichment of butyrate-producing bacteria in the intestinal tissue.
How might it impact on clinical practice in the foreseeable future?
▸ The CRC-associated microbiota is being continually defined as new biomarkers of CRC are discovered. The microbial dysbiosis observed in patients with CRC may be manipulated by probiotic bacteria, and the probiotic strains used in this study show promise as a beneficial component of treatment and therapeutic development in CRC.
Introduction
Colorectal cancer (CRC) currently affects ∼1.4 million people each year and its incidence is increasing worldwide.1 Despite novel treatment strategies, the mortality rate is very high among patients with advanced stages of the disease. The majority of CRC cases (70%) arise sporadically in a time-dependent manner according to the adenoma-carcinoma sequence of genetic alterations.2 Accumulating evidence points to a strong link between lifestyle factors and the risk of developing CRC. Risk factors include advanced age, tobacco and alcohol consumption, physical inactivity, increased body weight and diet (eg, high consumption of red and processed meat), with the latter being the most significant.3 The strong connection to lifestyle factors indicates that CRC incidence can be influenced by lifestyle changes.
Microbial imbalance (dysbiosis) in the gut can be caused by environmental factors (eg, diet, infection, antibiotics), but little is known about how the composition of the microbiota affects development of CRC. We hypothesise that the association between CRC risk and diet is partially mediated by the gut microbiota. Recent studies show that the gut microbiota differs between patients with and without CRC or colon adenomas, that is, precancerous lesions that may develop into CRC, and that the microbiota is a risk factor for cancer development.4 ,5 Increased bacterial diversity has been reported in the microbiota of patients with colon adenomas and tumours compared with non-CRC controls.6–8 The CRC-associated microbiota also has a microbial profile distinct from healthy tissue, including bacteria that thrive in the cancer-related microenvironment. The proliferation of carcinoma-associated taxa such as Fusobacterium in tumours is a potential microbial biomarker of a dysbiotic microbiota in CRC.8 ,9 This microbial dysbiosis may reduce the regulatory effect of commensal bacteria on cell proliferation in colon mucosa and contribute to the development of adenomas.10 It is plausible that dietary changes or intervention with probiotic bacteria may reduce the risk of CRC development; however, it is not known whether specific tumour-associated alterations in the microbiota are modifiable in patients who manifest disease.
Probiotics are defined as live microorganisms that, when administered in adequate amounts, confer a health benefit on the host.11 Studies have demonstrated beneficial effects of probiotic bacteria on reducing CRC tumour development and mucosal inflammation in animal models; however, supporting clinical data in humans are limited.12 To investigate the potential of altering the microbiota in patients with colon cancer, we conducted a prospective intervention study using selected probiotic strains. The bacterial strains used in this study, Lactobacillus acidophilus NCFM and Bifidobacterium animalis subsp. lactis Bl-04, have a long history of safe use as commercial probiotics and documented health benefits. NCFM has shown efficacy for colonic tumour growth attenuation in rodents and was associated with reduced levels of procarcinogenic metabolites in the human gut.13 ,14 Bl-04, although less studied with relation to CRC, has anti-inflammatory properties and was shown to alleviate colitis in mouse models.15 In this pilot study, we obtained intestinal tissue and faecal samples from patients with colon cancer that received or did not receive probiotics, and from non-cancer controls, to characterise the colon cancer-associated microbiota and determine whether this signature could be altered by probiotic intervention.
Patients and methods
Study outline
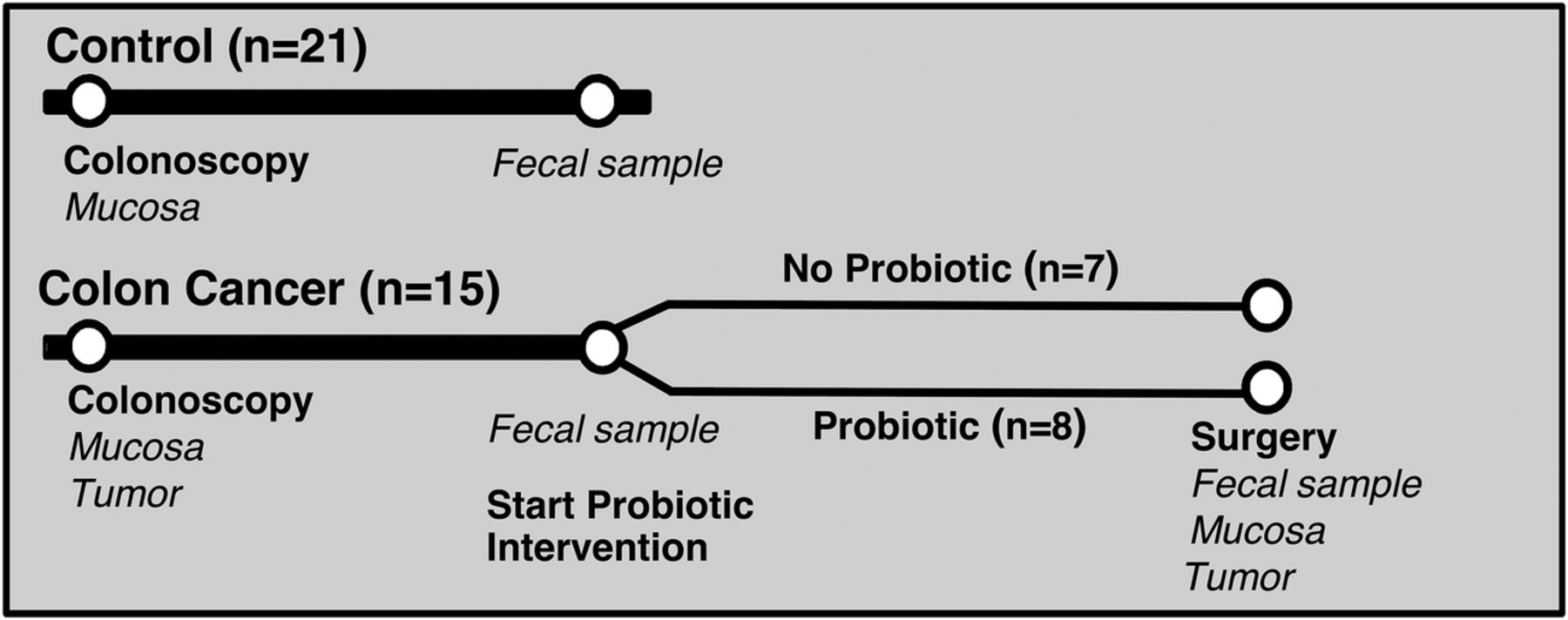
Samples were obtained from patients with colon cancer at colonoscopy and at surgery, and the colonic microbiota was studied in a prospective manner (figure 1). Patients without cancer or adenomas at colonoscopy were included as controls for comparisons of baseline data. In addition, a prospective randomised intervention with probiotics in patients with colon cancer was carried out (figure 1). The study was approved by the Regional Ethical Review Board in Gothenburg under study number 233-10 and registered at ClinicalTrials.gov (ID: NCT03072641). Informed consent was obtained from all study subjects.

Clinical study outline and sample collection for microbiota analyses.
Non-cancer controls
Twenty-one non-cancer controls were included in the study (table 1). Colonoscopy was performed due to abdominal symptoms such as diarrhoea, constipation, abdominal pain, or lower gastrointestinal bleeding and iron-deficient anaemia. The prerequisites for inclusion into the control group were a normal-appearing colonic mucosa. Study subjects with significant pathology such as colonic polyps, inflammatory bowel disease, malignancy or ischaemic colitis were excluded. Microscopic colitis was ruled out by light microscopic examination of biopsy specimens obtained from the mid-portion of the ascending and sigmoid colon. The presence of colonic diverticula was accepted provided there were no signs of acute diverticulitis or diverticulosis-associated colitis. Study subjects who received recent antibiotic therapy or consumed probiotics regularly were also excluded.
Study population characteristics
Colon cancer patients
Fifteen patients who were diagnosed with stage I–III colon cancer at colonoscopy were included in the study (table 1). Tumours were classified according to the tumour-node-metastasis (TNM) staging system.16 The prerequisite for inclusion into the colon cancer group was the presence of at least one malignant tumour in the colon and ≥18 years of age. Study subjects that had adenomas, received recent antibiotic therapy or consumed probiotics regularly were excluded.
Probiotic intervention
Patients with cancer were randomised to receive (n=8) or not to receive (n=7) probiotic supplementation. The intervention started at the second revisit to the clinic and continued until the day of surgery (average length of intervention was 31±28 days; range 8–78 days). The probiotic supplementation consisted of two ProBion Clinica (Wasa Medicals AB, Halmstad, Sweden) tablets, yielding a daily dose of 1.4×1010 CFUs Bifidobacterium lactis Bl-04 (ATCC SD5219), 7×109 CFUs Lactobacillus acidophilus NCFM (ATCC 700396) and 0.63 g inulin. Flow cytometry analysis showed that on average 85% of cells were live cells with non-compromised cell membranes. The tablets were produced using a low-compression technique which results in greater cell survival during production and prolonged stability compared with conventional techniques. In addition to a slow-release profile (150–180 min), the tablets improve survival of the probiotic bacteria in the gastrointestinal tract.
Tissue and faecal sample collection
In controls, tissue samples were obtained from the mid-portion of the ascending colon (right side samples) and the sigmoid colon (left side samples) using regular biopsy forceps. One of these samples was used for analysis. In patients with colon cancer, biopsies were taken during colonoscopy from the tumour as well as from normal-appearing mucosa distant from the tumour. There were no complications related to the colonoscopy or biopsy procedures. Faecal samples were obtained post colonoscopy and frozen at −80°C within 24 hours. All participants attended colonoscopy and provided a baseline faecal sample. Thereafter the patients with colon cancer were randomised to receive, or not receive, probiotics and eventually attended surgery. At surgery, tumour tissue, macroscopically normal mucosal samples obtained 10 cm from tumours, and faecal samples were obtained, snap-frozen in liquid nitrogen and stored at −80°C until analysis. Owing to healthcare procedures, the timing between colonoscopy and faecal sample donation or randomisation and surgery (ie, length of intervention) could not be controlled beforehand. Hereafter non-tumour biopsies taken from patients with colon cancer and control participants are referred to as mucosa and control samples, respectively.
DNA isolation and 16S rRNA gene sequencing
DNA was extracted from the samples with the Promega Wizard Genomic DNA Purification Kit (Promega Corporation, Madison, Wisconsin, USA) as described previously17 ,18 followed by PCR inhibitor removal with OneStep-96 PCR Inhibitor Removal Kit (Zymo Research, Irvine, California, USA), and DNA concentration measurement with Qubit 2.0 Fluorometer (Life Technologies, Darmstadt, Germany). The microbiota composition was analysed by targeted PCR amplification of the V4 variable region of the 16S rRNA gene of bacteria and archaea. Microbial DNA was amplified in triplicate PCR with primers 515F (5′-GTGCCAGCMGCCGCGGTAA) and 806R (5′-GGACTACHVGGGTWTCTAAT) as described.19 ,20 The PCR amplification proceeded at 95°C for 3 min for initial denaturation, followed by 30 cycles at 95°C for 45 s, 55°C for 60 s and 72°C for 90 s; and final extension of 10 min at 72°C. PCR products were purified and normalised using the SequalPrep Normalisation Plate Kit (Thermo Fisher Scientific, Waltham, Massachusetts, USA). Paired-end 250 bp reads were generated from amplicon libraries with the MiSeq instrument (Illumina, San Diego, California, USA).
Analysis of microbiota composition
Sequence analysis was conducted with the Quantitative Insights Into Microbial Ecology (QIIME V.1.9.1) bioinformatics pipeline.21 Reads were paired using fastq-join22 and those with a Phred quality score <20 were discarded. An open reference scheme with uclust23 was used for clustering reads into operational taxonomic units (OTUs) at 97% sequence similarity, such that OTUs not matching a reference sequence in the Greengenes database24 ,25 (V.13.8) were clustered de novo. Python nearest alignment space termination tool (PyNAST)26 was used for sequence alignment, and a taxonomic tree was constructed with FastTree-2.27 OTUs containing <5 sequences were removed. After quality filtering, 11 276 994 sequences were retained, with an average of 99 796 sequences per sample. Metrics for α-diversity (within-sample richness), including observed OTUs and phylogenetic diversity (PD),28 and β-diversity (pairwise dissimilarity) UniFrac distance29 were calculated on OTU tables rarefied to a depth of 10 695 sequences.
Statistics and network analyses
Group comparisons for diversity metrics were conducted within QIIME and graphed using Prism V.7 (GraphPad Software, La Jolla, California, USA). α-Diversity comparisons and group distances for β-diversity (weighted UniFrac metric) were generated with a non-parametric t-test using 1000 Monte Carlo permutations. Discriminate taxa (>0.1% abundance) between groups were identified with the Wilcoxon rank sum test in QIIME. Adjusted p values controlling the false discovery rate (FDR)30 are reported where appropriate.
A correlation network analysis was constructed for tumour samples using the CoNet software31 for genus-summarised abundance data and clinical factors of disease and tumour severity. Pairwise correlations (Pearson and Spearman), Bray Curtis dissimilarity and Kullback-Leibler divergence were used to create an initial association network. The edgeScores randomisation routine was used where row-wise permutations were calculated with 100 iterations and the 1000 highest and lowest scoring edges were retained. Renormalisation option was enabled. A second network was created with 100 bootstrap iterations and merged into one final network. The p values were merged using the Brown option and adjusted with the Benjamini-Hochberg FDR correction at a threshold of 5%. Only significant edges supported by a minimum of two methods were retained. The network was visualised in Cytoscape (V.3.1.1).
Heat maps were generated using a two-way hierarchical cluster analysis with Ward's minimum variance method. Data were standardised across individual groups by Z-scores, where group abundance was subtracted from the population mean and divided by the SD in JMP V.9 (SAS Institute Cary, North Carolina, USA).
Results
Microbiota signature of colon cancer
Diversity is increased in the colon cancer microenvironment
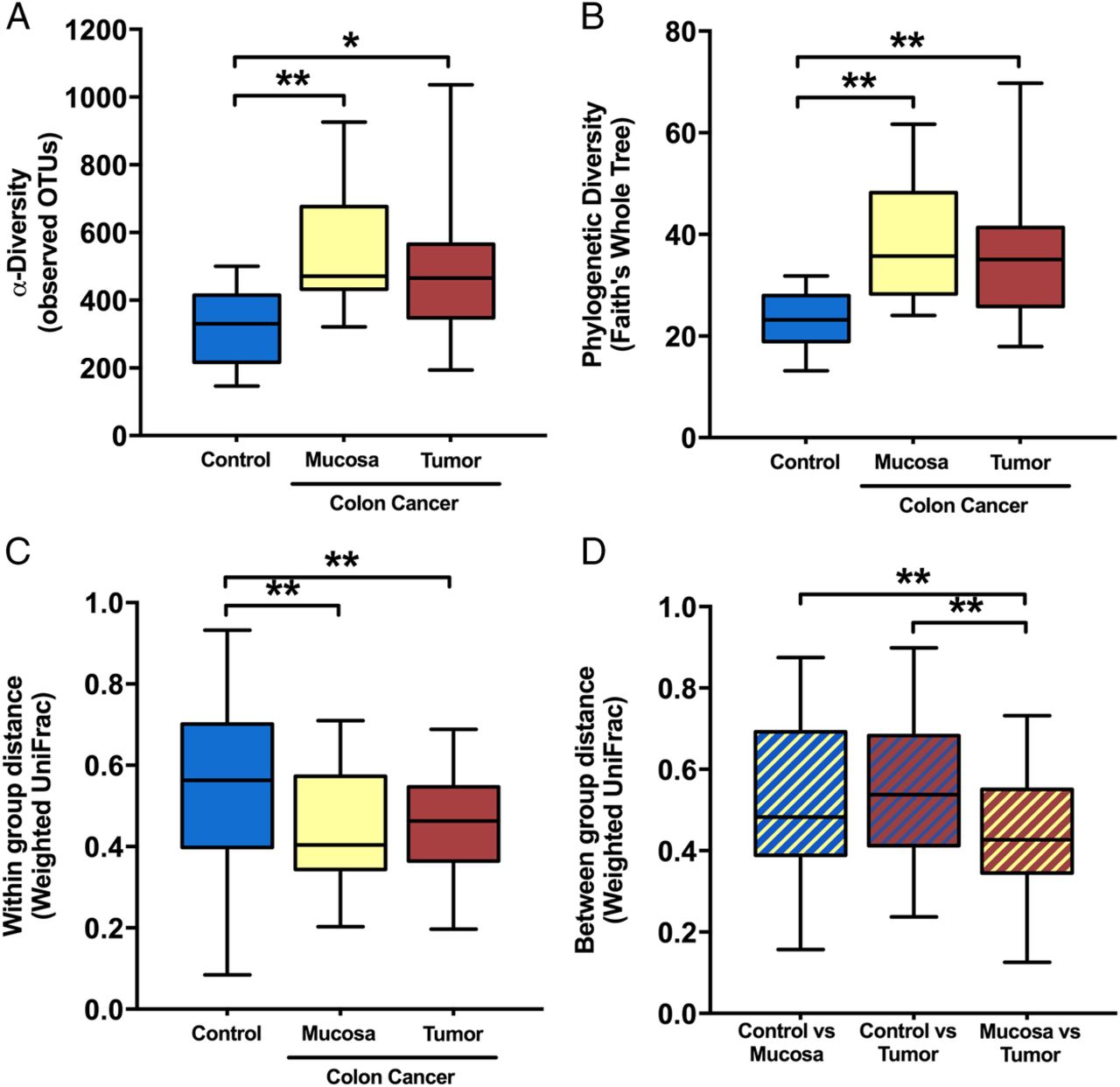
Microbial diversity was significantly enriched in the colon cancer mucosal microbiota. α-Diversity, as measured by the number of observed OTUs (figure 2A) and Faith's PD Whole Tree index (figure 2B), was increased in both tumour and mucosa samples from patients with colon cancer compared with control samples (FDR≤0.05); however, no significant differences were found between groups for faecal samples (data not shown; FDR>0.1). β-Diversity based on the weighted UniFrac distance revealed that the tissue microbiota was more similar among patients with colon cancer within tumour or mucosa biopsies than among control samples (figure 2C). The microbiota composition from the mucosa adjacent to the tumour was more similar to the tumour-associated microbiota than to that of control samples (figure 2D). For patients with colon cancer, α-diversity or β-diversity did not differ between the tumour-associated microbiota and the macroscopically healthy appearing mucosa-associated microbiota sampled from the same patients at a distance of 10 cm from the tumour (FDR>0.1).
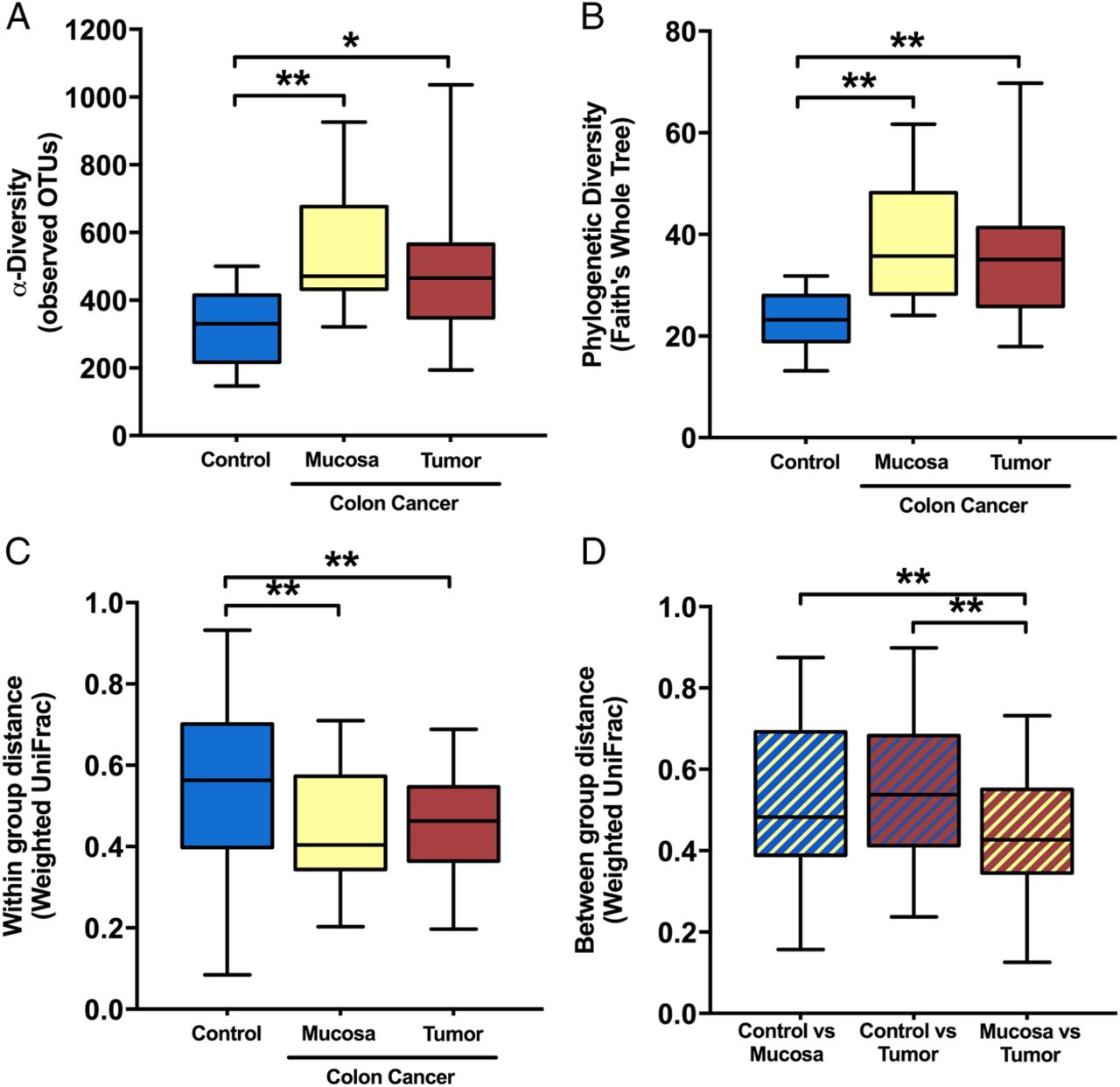
α-Diversity metrics: (A) observed operational taxonomic units (OTUs) and (B) phylogenetic diversity (PD whole tree) were greater in the mucosa and tumour microbiota of patients with colon cancer compared with the non-cancer control mucosa at colonoscopy. β-diversity: (C) within-group weighted UniFrac dissimilarity) was lower for colon cancer mucosa or tumour microbiota than control mucosa and (D) between-group dissimilarity was lowest for colon cancer mucosa versus tumour samples when compared with controls at colonoscopy. False discovery rate (FDR)-corrected non-parametric t-test using 1000 Monte Carlo permutations; **FDR<0.01; *FDR<0.05.

Microbial interaction network for (A) colon cancer tumour microbiota and (B) subset of direct connections to colon cancer-associated taxa in tumour samples. Nodes represent genus-level summarised taxa and are coloured by phylum. Edges were retained only if supported by two of the following methods: Spearman, Pearson, Bray-Curtis and Kullback-Leibler and FDR≤0.05. Positive associations are denoted by green edges and negative associations are denoted by red edges. FDR, false discovery rate.
Differentially abundant taxa in patients with colon cancer compared with non-cancer controls
The overall composition of the microbiota was altered in the patients with colon cancer compared with controls, and the abundances of several taxa were elevated in samples obtained from the tumour microenvironment. The phylum Fusobacteria and genus Fusobacterium were highly enriched in colon cancer samples (FDR≤0.05), where the mean abundance was >7% in the tumour tissue and <0.5% in control samples (FDR≤0.05; see online supplementary table S1 and table 2). The phylum Euryarchaeota and genus Methanobrevibacter within were also enriched in colon cancer samples (FDR≤0.05; see online supplementary table S1 and table 2). Several changes in faecal and mucosal microbial composition were due to genera within the phylum Firmicutes, despite there being no overall difference at phylum level (table 2, see online supplementary tables S1 and S2). Clostridium and Dialister were among the genera enriched in the tumour and mucosa samples from patients with colon cancer (FDR≤0.05; table 2). Peptostreptococcus was significantly more prevalent in all sample types derived from patients with colon cancer (mucosa, tumour and faeces) (FDR≤0.05; see online supplementary table S3 and table 2). Unclassified Clostridiales and Oscillospira were elevated in the mucosa and Selenomonas in the tumour from patients with colon cancer (FDR≤0.05; table 2). Rikenellaceae spp (phylum Bacteroidetes) and Bilophila (phylum Proteobacteria) were also greater in the mucosa from patients with colon cancer, while Streptococcus was depleted (FDR≤0.05, table 2). When faecal samples from the controls and patients with colon cancer post colonoscopy (no intestinal cleansing procedure included) were compared, Clostridiaceae spp and Dorea, in addition to Peptostreptococcus, were more abundant in the colon cancer group while Tenericutes (phylum) and Roseburia were reduced (FDR≤0.05; see online supplementary table S2).
supplementary data
Microbiota alterations in colon cancer mucosa and tumour samples compared with non-cancer control mucosa at colonoscopy
Microbial interactions in colon cancer tumour tissue
To explore the complex microbial interactions in the tumour tissue, we constructed a correlation network to identify patterns of co-occurring microbes. The resulting network contained 61 nodes and 350 significant edges between microbial genera (figure 3A). Based on the clustering patterns observed, groups of co-occurring taxa within the tumour microbiota were evident. Subsetting the edges that corresponded to tumour-enriched genera showed that several of these taxa co-occurred within the same samples (figure 3B). Fusobacterium, the most over-represented genus in tumour samples, tended to co-occur with Peptostreptococcus, Campylobacter and Bulleidia. Peptostreptococcus and Selenomonas, also highly elevated in tumour samples, were positively associated with each other, as well as with Parvimonas and Mogibacteriaceae spp. Conversely, tumour-associated Methanobrevibacter was negatively correlated with both Fusobacterium and Selenomonas. Correlation with host parameters of disease or tumour severity did not reveal any substantive associations with specific taxa, possibly due to the lack of dichotomy within our cohort for these clinical factors (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Heat map with two-way hierarchical clustering of genus-summarised microbiota abundance and sample grouping for mucosal and tumour microbiotas from patients with colon cancer at colonoscopy and surgery (with or without probiotics), and for non-cancer control mucosal microbiota at colonoscopy.
Probiotic intervention alters the colon cancer microbiota
Microbiota composition shifts with probiotic intervention
Cluster analysis revealed that the overall composition of the microbiota in the mucosa and tumour samples from patients with colon cancer that consumed the probiotic differed distinctly from patients with cancer who did not receive the probiotic, as well as from the non-cancer control patients (figure 4). The mucosa from the probiotic-supplemented patients with cancer was similar in composition to the tumour tissue at surgery. A large cluster containing several butyrate-producing bacteria from the phylum Firmicutes was apparent in the patients that received the probiotic, and was distinctly enriched compared with the colon cancer patients that did not receive the probiotic and control patients.
Butyrate-producing bacteria are enriched with probiotic intervention
The overall abundance of the phylum Firmicutes was significantly increased at the time of surgery in faecal samples from patients with colon cancer that received probiotic intervention compared with those that did not (77% vs 63%; FDR≤0.05) (see online supplementary table S4). Within the Clostridiales, several butyrate-producing genera were consistently elevated in the different sample types from the patients that received the probiotic (table 3). Clostridiales spp and Faecalibacterium were enriched in all sample types obtained from patients that received the probiotic (p≤0.05, FDR not significant). Eubacterium was elevated in faecal and mucosa samples, and Roseburia and Lachnospira were greater in mucosa and tumour samples in patients that received the probiotic (p≤0.05, FDR not significant). The CRC-associated taxa, Fusobacterium and Peptostreptococcus, were less abundant in patients that received the probiotic, but this was only detected in faecal samples (p≤0.05, FDR not significant) (table 3). For bacteria that were enriched in the patients with colon cancer that received probiotics, we did not find significant correlations between bacterial abundance and length of probiotic intervention; however, the power of detection may have been limited by small sample size (data not shown, Spearman correlation p>0.1).
Microbiota alterations with probiotic intervention in patients with colon cancer at surgery
Discussion
Interactions between the intestinal microbiota and CRC may be bidirectional, where a dysbiotic microbiome promotes carcinogenesis, or possibly the tumour microenvironment and related inflammatory state cause microbial dysbiosis.32 ,33 Our results establish that a distinct microbial community exists in the colonic tumour and adjacent mucosa, which was characterised in our study by increased microbial diversity and the differential abundance of numerous bacterial groups compared with non-cancer control samples. A corresponding differential clustering of CRC-associated mucosal microbiotas from those of healthy volunteers has been reported in several 16S rRNA-based sequencing analyses, although it was not always linked to increased α-diversity.8 ,32 ,34 ,35 The enrichment of microbial diversity in CRC tumour tissue contrasts with trends of greater diversity typically observed in microbiota samples derived from healthy populations. Putative explanations for the increased diversity in CRC tumour tissue are that the nutrient-rich microenvironment may support a more diverse consortium of microbial species8 or that it represents a transient state during dysbiosis formation. This finding is noteworthy considering that the increase in diversity was not observed in colon cancer-derived faecal samples, and therefore may signify that this feature is characteristic of the tumour microenvironment.
The colon cancer-associated microbiota signature in our study was characterised in part by increased abundance of Peptostreptococcus and Fusobacterium, both genera that contain opportunistic oral pathogens known to cause infections such as periodontitis. The over-representation of these typically oral-associated organisms in CRC-associated samples is especially intriguing and has been recently examined in the literature.36 ,37 Fusobacterium has several attributes that make it uniquely adapted to the tumour-microenvironment, such as being asaccharolytic, and it has been shown to accelerate tumorigenesis and promote a pro-inflammatory environment.37 ,38 These organisms have been repeatedly associated with CRC-associated microbiotas and therefore have been proposed as microbial markers for CRC.9 ,32 ,39 The consistency of data linking these specific genera with CRC is substantial, considering the complexity of the gut microbiome and differences in population attributes, sampling and analyses methods across studies. Specifically, Fusobacterium nucleatum, Peptostreptococcus anaerobius and Parvimonas micra have shown promise as microbial CRC markers due to an increased relative abundance of 132-fold, 37-fold and 41-fold, respectively, among patients with CRC in a large cohort study (103 CRC patients and 102 controls analysed).9 We found that Peptostreptococcus was increased in all three colon cancer-associated sample types (mucosa, tumour, faeces) and was not present at detectible levels in non-cancer control samples. Similarly, Fusobacterium was elevated over 30-fold between tumour and control biopsies, supporting the importance of these bacteria as potential CRC markers. Additionally, we have shown that these CRC tumour-associated organisms are positively associated and co-occur within the same samples. Their known high capacity to colonise the mucosa and form biofilms in the oral microbiome may suggest that a similar mechanism of colonisation occurs in CRC. Oral commensals and pathogens that are associated with CRC may act as dysbiosis-triggering organisms,33 and further studies are warranted to determine whether the origin of specific strains can be linked between oral and CRC tumour sites.40
Additional differentially abundant taxa over-represented in colon cancer tumour and mucosa samples included the phyla Tenericutes and Euryarchaeota as well as the genus Methanobrevibacter. Tenericutes was found to be less abundant compared with controls in faecal samples suggesting the colon cancer association was localised to the mucosal surface or that different organisms were detected from this phylum between the sample types. Both Tenericutes and Methanobrevibacter have previously been associated with adenoma or CRC microbiotas, and the phylum Tenericutes includes parasitic pathogens (Mollicutes) that have previously been suspected as causal agents in other cancers.8 ,34 ,41 Methanobrevibacter has been linked to a multitude of intestinal disorders, periodontitis and CRC, although the mechanism of CRC association is unknown.42 It was negatively associated with Fusobacterium in our network analysis, suggesting a different mechanism of involvement in CRC. Other tumour-associated genera were from within Firmicutes (Selenomonas, Clostridium, Dialister and Parvimonas); however, contradictory findings have been published on their presence in adenoma and carcinoma-associated tissue.5 ,34 The non-cancer controls displayed the greatest amount of within-group variability in microbiota composition; however, the increased abundance of genus Streptococcus was evident in control participants' biopsies relative to colon cancer. Streptococcus bovis has previously been associated with CRC tumours,32 ,35 but in our data this elevation appears to be primarily attributed to sequences related to Streptococcus thermophilus. It could be interesting to further investigate whether S. thermophilus may have protective properties in healthy populations or if it is solely depleted in patients with colon cancer. The control patients were not diagnosed with colon cancer; however, colonoscopy was performed due to various gastrointestinal complaints with manifest symptoms, which may partially explain the high level of variability among their microbiota profiles.
Overall, the composition of the microbiota in samples from patients with colon cancer that received probiotics had a unique profile characterised by an increased abundance of butyrate-producing bacteria in tumour, mucosa and faecal samples compared with patients with cancer who did not receive probiotics. Butyrate interacts intimately with colonic epithelial cells as an energy source for colonocytes and by modulating signalling pathways. It plays a beneficial role in colon cancer by inhibiting cell proliferation, reducing IFN-γ-mediated inflammation and promoting cell apoptosis and tumour suppressor gene expression.43–45 Clostridiales spp and butyrate-producing Faecalibacterium, Roseburia and Eubacterium were enriched in samples obtained from patients with colon cancer with probiotic intervention. Despite the FDR-corrected p values not reaching statistical significance, this finding was detected consistently in tumour, mucosa and faecal samples. A depletion of butyrate-producing bacteria in the microbiota has been reported in patients with various stages of CRC progression,46–48 and butyrate's tumour-suppressive properties have been shown to be directly mediated by the gut microbiota, further supporting its importance in CRC.49 Additionally, the faecal microbiota of patients with colon cancer taking probiotics had reduced levels of CRC-associated genera Fusobacterium and Peptostreptococcus according to the non-FDR-corrected p value. This finding is in accordance with a previous probiotic intervention trial where supplementation with Bifidobacterium longum, Lactobacillus acidophilus and Enterococcus faecalis reduced Fusobacterium and Peptostreptococcus in CRC patients to a level comparable to healthy controls.39 As these two genera are strongly associated with CRC microbiota in several studies, these results emphasise the value of evaluating probiotics for CRC prevention and care. Moreover, probiotics have also been shown to mediate inflammatory responses, as Gianotti and colleagues observed that the mucosal colonisation of probiotic strain Lactobacillus johnsonii La1 was correlated with reduced proliferation and modulation of specific dendritic cells in CRC.50 Unfortunately, we were unable to achieve the level of sensitivity necessary to differentially detect colonisation of our specific probiotic strains from the native populations by qPCR in the mucosal samples from this study (data not shown).
The difficulty in obtaining intestinal mucosal and tumour samples as compared with faecal sampling for microbiota analysis presented challenges in the study design and sampling. Patients underwent bowel cleansing prior to colonoscopy but no bowel preparation was done prior to surgery, and the timing of the faecal sample collection post colonoscopy was not controlled, both of which may influence the microbiota profiles. Additionally, the intervention length varied among patients as it would have been unethical to restrict the time to their surgery. We therefore chose to focus primarily on comparisons among samples that were obtained either at the time of colonoscopy or at surgery. In a future study, it would be preferable to more stringently control participant groups, but we avoided this in the pilot trial primarily for ethical reasons. Despite these limitations, by obtaining tumoral, mucosal and faecal samples, we assessed the colon cancer-associated microbiota by several comparisons: (1) tumour to mucosa within close proximity; (2) the aforementioned to non-cancer control samples; (3) colon cancer faeces to non-cancer control faeces; (4) colon cancer probiotic intervention to no probiotic intervention. Fewer differentially abundant taxa were detected in the faecal microbiota than the corresponding tissue microbiota, suggesting that the tissue microbiota profile is more informative for identifying putative microbial markers of colon cancer. Bacterial adherence to the intestinal epithelium or biofilm formation may have contributed to the differences we observed between the tissue and faecal samples, but tissue samples more likely represent organisms that directly interact with host and immune cells and are thus preferable to more easily obtained faecal samples. Peptostreptococcus, however, was significantly enriched in both tissue and faecal samples and shows promise as a microbial CRC marker.
The results of this study support the hypothesis that the colon cancer-associated microbiota can be manipulated by specific probiotic strains, resulting in an altered microbiota enriched with beneficial bacteria. Our study provides evidence that microbiota modulation by probiotics could be considered as part of a therapeutic regime for CRC patients. Further studies should be conducted in a larger population to confirm these initial findings, and ideally should be complemented with metabolomics data to elucidate the role of butyrate. Future studies could also expand beyond the microbiota to include the potential influences of fungi (mycobiome), viruses (virome) and microbial bioactive molecules on CRC development. The interplay between diet, microbiota and host in maintaining homeostasis is an important consideration in therapeutic strategies for CRC, and analysis of the microbiome is a critical component in understanding how these complex interactions influence the development and progression of carcinogenesis.
Acknowledgments
The authors thank Ann-Louise Helminen and Hillevi Björkqvist for inclusion of study subjects and collection of samples, Lena Munro for work with the clinical database and Jaqueline Flach, Marianne Åkerström, and Elisabeth Odin for handling of samples. The authors also thank Nicolas Yeung, Krista Salli, Paige Roos and Wes Morovic for technical work in preparing the samples for sequencing and Buffy Stahl for supporting project management.
References
Footnotes
Contributors YW, PR, AL, ACO and HL designed and executed the clinical research study. YW was the principal investigator. AAH analysed the data and performed statistics. AAH, AL, ACO and YW interpreted the data. AL and YW supported data integration and project management. LC contributed expertise on ProBion Clinica. AAH, AL and YW wrote the manuscript. All authors reviewed and approved the final manuscript.
Funding This work was supported by a grant from the Swedish state under the LUA/ALF agreement (grant number ALFGBG-542821).
Competing interests At the time of the research, AAH, AL and ACO were employees of DuPont, who manufactures the probiotic strains used, and LC is an employee of Wasa Medicals who produced the probiotic product used in this study.
Patient consent Obtained.
Ethics approval The study was approved by the Regional Ethical Review Board in Gothenburg under study number 233-10.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement No additional data are available.