Article Text

Abstract
Introduction Spontaneous bacterial peritonitis (SBP) is mainly caused by bacterial translocation of enteric Gram-negative bacteria, predominantly Escherichia coli. The sympathetic nervous system (SNS) is activated in advanced cirrhosis, particularly in the splanchic circulation, and exerts potent immunosuppressive actions. However, the role of splanchnic SNS activity in bacterial translocation and bacterial spreading in cirrhosis remains unclear.
Methods E coli or Stapylococcus aureus (106 CFU) were given intraperitoneally. After 6 h, mesenteric lymph nodes (MLN), liver, spleen, lung and peripheral blood were harvested from ascitic cirrhotic rats (LC) and healthy controls with and without splanchnic sympathectomy (SE). The bacterial tissue burden was determined by standard microbiological culture techniques. In vitro phagocytic activity of peritoneal polymorphonuclear leucocytes was assessed by FACS analysis.
Results Under basal conditions SE reduced bacterial translocation to MLN in LC rats from 45% to 17%. LC rats had a marked increase in bacteraemia after E coli and S aureus challenge and an increased incidence and degree of E coli translocation to MLN, liver, spleen and lung compared with control rats. SE prevented bacteraemia in LC rats after E coli but not after S aureus challenge. Prior SE abolished the difference in incidence as well as the bacterial tissue burden in each organ after E coli application in LC rats, being no longer significantly different from control rats with or without SE. The protective effects of SE against E coli were associated with a greater influx of mononuclear cells into the peritoneal cavity and increased phagocytic activity of peritoneal polymorphonuclear leucocytes.
Conclusions In cirrhosis with bacterial peritonitis, hyperactivity of the splanchnic sympathetic nervous system contributes to the translocation of E coli but not S aureus to MLN and extraintestinal sites. This indicates a key role for sympathetic drive in the impairment in host defence against Gram-negative bacteria in cirrhosis.
- Sympathetic nervous system
- bacterial translocation
- spontaneous bacterial peritonitis
- liver cirrhosis
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
Spontaneous bacterial peritonitis (SBP) is defined as polymorphonuclear (PMN) cell count in ascitic fluid ≥250/mm3 in absence of an intra-abdominal source of infection or malignancy.
Norepineprine, dependent on local concentration and target tissue, can exert potent immunosuppressive actions including chemotaxis.
E coli is one of the most frequently isolated bacteria in SBP and/or bacteraemia in cirrhosis, a phenomenon attributed to pathological bacterial translocation from the gut.
Non-selective β blockers are recommended for prophylaxis of variceal haemorrhage in portal hypertension, but their impact on the development and course of Gram-negative infections such as E coli in cirrhosis has not been investigated systematically.
What are the new findings?
Influx of PMN cells into the peritoneal cavity in cirrhotic ascitic rats depends on the type of bacteria.
Splanchnic sympathectomy increases the influx of PMN cells into the peritoneal cavity in response to intraperitoneal E coli but not S aureus.
Splanchnic sympathectomy prevents spontaneous bacterial translocation from the gut to mesenteric lymph nodes and decreases the incidence and severity of systemic spreading of E coli (but not S aureus) including bacteraemia after its intraperitoneal application in ascitic cirrhotic rats.
How might it impact on clinical practice in the foreseeable future?
PMN cell count in ascites may be influenced by use of sympathetic nervous system (SNS)-modulating agents (eg, non-selective β blockers) and/or type of stimulating bacteria; this should be considered in respect to the definition and diagnosis of SBP.
Use of SNS-inhibiting agents such as non-selective β blockers could represent a new option for preventing E coli (or potentially other Gram-negative) infections in advanced cirrhosis.
Introduction
In cirrhosis of the liver, susceptibility to bacterial infections is greatly enhanced and the development of infectious complications is often fatal.1 2 Spontaneous bacterial peritonitis (SBP) is one of the most frequent bacterial infections in patients with advanced cirrhosis, occurring in the absence of any intra-abdominal treatable source of infection. The first episode of SBP is associated with mortality rates up to 40%, mainly due to secondary development of further complications such as hepatorenal syndrome, cardiocirculatory failure, variceal haemorrhage or hepatic encephalopathy.3 4 Moreover, bacteraemia develops 10 times more frequently in patients with cirrhosis than in the general population and represents an independent prognostic factor for a poor outcome in SBP.5 6 Bacteria isolated from ascites and/or blood during SBP are mainly Gram-negative enteral bacilli, with Escherichia coli representing the most common aetiological organism.6–8 However, an increasing incidence of SBP caused by Gram-positive bacteria in patients with decompensated cirrhosis has been observed.9 This has been attributed to a more frequent use of prophylactic antibiotic treatment and invasive procedures during hospitalisation.10
Bacterial translocation to mesenteric lymph nodes (MLN) is the pathophysiological hallmark for the development of SBP in liver cirrhosis and has been attributed to bacterial overgrowth, increased intestinal permeability and impaired host defence.11 Well-accepted risk factors predisposing to bacterial translocation and spontaneous bacterial infections in liver cirrhosis are advanced stage of disease and gastrointestinal haemorrhage.2 11 12 Polymorphonucelar leucocytes (PMNL) are the first line of host defence against bacterial infection and phagocytosis is one of their key capacities in order to kill invading microorganisms. Decreased phagocytic activity of PMNL, whether inherited or acquired in different chronic or acute diseases, increases the incidence and morbidity of infections.13–15 Several investigations have indicated that the phagocytic function of PMNL is impaired in liver cirrhosis.16–18 However, few studies have focused on the potent immunosuppressive action of the sympathetic nervous system, including inhibition of chemotaxis19 and the phagocytic activity of PMNL.20 21 These activities are essential for defence against Gram-negative bacteria. We recently reported that sympathectomy increases the influx of PMNL into the peritoneal cavity and enhances chemotaxis and the phagocytic capacity of peritoneal PMNL, thereby preventing the systemic spread of E coli from the peritoneal cavity in healthy mice.22 Sympathetic hyperactivity is a well-known phenomenon that develops in cirrhosis; it is most marked in end-stage disease with ascites23 24 and is particularly excessive in the hepatosplanchnic circulation.25 26
We therefore hypothesised that the well-known splanchnic sympathetic hyperactivity in liver cirrhosis with ascites has an unfavourable effect on clearance of E coli in an animal model of SBP. We applied a newly developed method of specific splanchnic sympathectomy (SE) to ascitic cirrhotic rats to investigate the effect on (1) the development of bacterial translocation to MLN; (2) the spread of E coli or S aureus from the peritoneal cavity to extraintestinal sites in cirrhosis with ascites; and (3) the peritoneal influx and phagocytic activity of PMNL in response to E coli.
Methods
Animal models
Cirrhosis was induced in male pathogen-free CD rats (Charles River, 50–80 g initial weight) by inhalation of carbon tetrachloride (CCl4) together with phenobarbital (0.35 g/l) in the drinking water, as previously described.27 28 CCl4 administration was started three times a week over 1 min and increased every other week by 1 min to a maximum of 5 min, depending on the change in the body weight of the animal. After 12–16 weeks this approach induces micronodular liver cirrhosis with ascites. Seven days before the experimental procedures were undertaken the application of CCl4 and of phenobarbital was stopped. Only cirrhotic animals with decompensation of liver function and thus presence of ascites were used. Phenobarbital-treated age- and sex-matched rats were used as a control group.
Splanchnic sympathetic denervation
In order to focus on the role of the splanchnic sympathetic nervous system, we developed a method of exclusive chemical sympathectomy (SE) in the splanchnic and peritoneal area. This is necessary to exclude an influence of extraintestinal and extraperitoneal sympathetic nerves on the results obtained. In particular, central sympathetic nervous system efferent functions are known to influence systemic and hepatosplanchnic vascular derangement in cirrhosis.29 Furthermore, cardiac and renal adrenergic innervation is key in vascular and plasma volume homeostasis in severe portal hypertension.23 A specific splanchnic SE was therefore achieved by utilisation of saporin-coupled anti-dopamine-β-hydroxylase (Anti-DBH, 5 μg/ml intraperitoneally in 3 ml sterile 0.9% saline, Cat. Nr. IT-03, Advanced Targeting Systems, San Diego, California, USA). After binding to DBH and endocytosis, this immunotoxin is transported retrograde to the nucleus of the axon30 where saporin is released.31 Saporin irreversibly blocks protein synthesis, inducing cell death.32 Application was started in cirrhotic animals at the time of first appearance of ascites, ensuring an advanced and irreversible stage of liver cirrhosis. Healthy age-matched control rats were injected in parallel. Anti-DBH was dissolved in sterile saline and injected intraperitoneally 3 weeks before starting the experimental procedures (see below). Saline alone was given to healthy control and cirrhotic rats serving as control groups. The efficacy of SE was assessed by immunohistological analysis of the spleen and jejunum (representing splanchic tissue) and the heart (representing extraintestinal sites).
Immunohistochemistry of sympathetic innervation of small intestine
Samples of small intestine from healthy control rats with and without SE (given 3 ml sterile 0.9% saline intraperitoneally as placebo) were fixed for 12–24 h in phosphate buffered saline (PBS) containing 3.7% formaldehyde and then incubated in PBS with 20% sucrose for 12–24 h.33 The tissue was then embedded in Tissue Tek and quick frozen. All tissue was stored at −80°C. Formaldehyde/sucrose-fixed tissue samples were cut into 5 μm thick sections. Immunohistochemistry was performed with a primary antibody against tyrosine hydroxylase (TH, the key enzyme for norepinephrine production in sympathetic nerve endings, cat. no. AB152, Chemicon, Temecula, California, USA). An Alexa 546 conjugated secondary antibody (cat. no. A-121010 against rabbit IgG, Molecular Probes, Leiden, The Netherlands) was used for immunofluorescent staining of sympathetic nerve fibres. For determination of intestinal innervation, 6–8 cryosections (5 mm thick) of the formaldehyde/sucrose-fixed tissue samples were used. The numbers of TH+ sympathetic single nerve fibres per square millimetre were determined by averaging the number of stained nerve fibres (minimum length 50 mm determined through a micrometre eyepiece) in 17 randomly selected high power fields of view (6400) as previously described.34 Positive nerve fibre staining was controlled by incubating the tissue with polyclonal control antibodies which always yielded a negative result. We investigated single nerve fibres because they reflect the neuronal innervation of non-neuronal target cells; we did not investigate neuronal plexus elements in the different layers of the intestine. In our hands, for single nerve fibre determination, automatic or semiautomatic image analysis is not feasible.
Experimental protocols
Protocol I assessed the impact of splanchnic SE on the incidence of bacterial translocation to MLN and rate of SBP in cirrhotic ascitic rats (n=23). Cirrhotic ascitic rats with and without prior SE (n=12 and n=11, respectively) were evaluated by microbiology as described below. Protocols II and III assessed the role of splanchnic SE in the spread of E coli and Staphylococcus aureus (S aureus), respectively, from the peritoneal cavity to intestinal and extraintestinal sites and peripheral blood. A modified experimental model of bacterial peritonitis was used35 in cirrhotic rats with and without prior SE (n=19 in each group). Age- and sex-matched healthy control rats with and without prior SE (n=19 in each group) were treated in the same manner. E coli and S aureus were grown to the mid-logarithmic phase in Luria broth with ampicillin. In a subgroup of animals, E coli was marked with green fluorescent protein (GFP). GFP-E coli was obtained by transformation of a clinical isolate of E coli with high-copy plasmid pCU18-GFP which carries a modified GFP gene.36 Cell density was estimated from the absorbance at 620 nm by use of calibration curves. Rats were injected intraperitoneally with 106 E coli or S aureus, respectively, suspended in 3 ml sterile saline. In preliminary experiments using E coli, these amounts led to a reproducible but modest degree of translocation to MLN, spleen and liver but almost negligible systemic spreading to the lung and no translocation to peripheral blood in control rats. Moreover, doses ≤107 CFU of E coli have been reported to be associated with no increase in mortality.35 Bacterial spreading was assessed after 6 h using standard microbiological techniques as described below.
Microbiological assessment
All experiments were performed under strict sterile conditions. Anaesthesia was induced with ketamine hydrochloride (Ketalar, 100 mg/kg body weight). Rats were shaved and the skin was disinfected with alcohol. Ascitic rats underwent paracentesis and samples of ascites were inoculated in aerobic and anaerobic blood culture bottles and a minimum of 1 ml stored at −80°C for later analysis. After midline laparotomy, MLN draining lymph nodes from the terminal ileum, cecum and ascending colon were dissected, removed and weighed. Tissue samples of liver, spleen and lung were also removed and weighed. MLN, liver, spleen and lung specimen were diluted in PBS (0.1 ml per 0.1 g), homogenised and 100 μl of suspension were cultured on McConkey, Mueller Hinton and Whole Blood agar for 48 h. Any growth of bacteria was considered as evidence of bacterial translocation. Homogenates were stored at −80°C for later analysis. Blood samples were obtained from the abdominal aorta and collected into heparin-containing ice-chilled tubes (stored at −80°C for later analysis) and fractions inoculated in aerobic and anaerobic blood cultures for 7 days. Any positive blood culture was regarded as evidence of bacteraemia. SBP was defined as any positive culture result from ascitic fluid (protocol I). Cell influx into the peritoneal cavity was assessed by standard differential blood count.
Histological analysis of the spleen, liver and lung
The spleen, liver and lung were removed 6 h after the initiation of infection and embedded in Tissue-Tek (Sakura Finetek, Europe) and quickly frozen by floating on liquid nitrogen. The tissue samples were cut into 4–6 μm thick sections and placed on precoated slides (SuperFrost Plus; Menzel-Gläser). To visualise nuclei, fixed samples were incubated with 4′6-diamidino-2-phenylindole hydrochloride (Sigma).
In vitro phagocytosis of E coli by PMNL
The phagocytic activity of PMNL from peritoneal fluid was determined in a separate protocol assessing cirrhotic rats with and without prior SE (n=5 and n=8, respectively) as well as control rats with and without prior SE (n=5 and n=9, respectively). In cirrhotic rats, ascites was obtained by total paracentesis whereas, in control rats, the peritoneal cavity was flushed with 30 ml isotonic sterile saline. The peritoneal fluid was centrifuged (10 min at 3000 rpm) and cell pellets were resuspended in FCS. Phagocytic activity was assessed by a commercial kit Phagotest (ORPEGEN Pharma, Heidelberg, Germany) according to the manufacturer's instructions as previously described.37 38 Briefly, cells were incubated in an ice bath for 10 min and 20 μl precooled FITC-labelled E coli bacteria (E coli, 2×107) opsonised with immunoglobulin and complement of pooled sera were added and incubated at 37°C for 10 min. A control (baseline) was kept on ice. The phagocytic process was stopped on ice and 100 μl ice-cold quenching solution was added to each probe to quench the FITC fluorescence of bacteria bound to the cell surface leaving the fluorescence of the internalised bacteria unaltered. After two washing steps (5 min, 250 g, 4°C), each probe was incubated at room temperature for 20 min. After addition of 2 ml of lysing solution provided in the kit for removal of erythrocytes and fixation of phagocytes. After a final washing step, the probes were incubated with 200 μl DNA staining solution at 0°C for 10 min. Cell suspensions were then measured by flow cytometry (10 000 events) within 60 min. PMNL were identified by flow cytometry (FACSCalibur, Becton Dickinson, Heidelberg, Germany) as described previously.39 The green fluorescence (FL1) of PMNL induced by FITC and the red fluorescence (FL2) of the PI-stained DNA of dead cells were analysed simultaneously with the cellular FSC/SSC with an argon laser tuned to 488 nm. PMNL were considered as active in the phagocytic process and oxygen radical production if FL1 was >101. Thus, active cells within the PMNL gate were determined by a marker M(a). The phagocytic activity (FL1active) was determined as mean FL1×M(a) (%). To allow comparison of individual results, the phagocytosis index was calculated as the ratio of fluorescence upon phagocytosis and fluorescence at baseline (FL1active stim/FL1active baseline). While the product of cells (%) participating in the phagocytic process and their mean fluorescence (FL1) determines the individual phagocytic capacity, the number of fluorescent cells (%) itself as determined by M(a) indicates the individual readiness of PMNL to participate in the phagocytic process. In order to assess this readiness of PMNL, we also compared the percentage of actively phagocytosing cells within the PMNL gate of each study group.
Statistical analysis
The results are expressed as mean±SE. Statistical analysis was performed using the Student t test for unpaired data using SPSS Version 13.0. The statistical significance level was p<0.05.
Results
Animal data
No difference in body weight, weight of MLN or lung was observed between the study groups (table 1). However, cirrhotic rats with and without SE had increased spleen and liver weight compared with control groups. No difference in weight of spleen, liver or MLN was detectable between cirrhotic rats with and without SE.
Animal data
Splanchnic sympathectomy
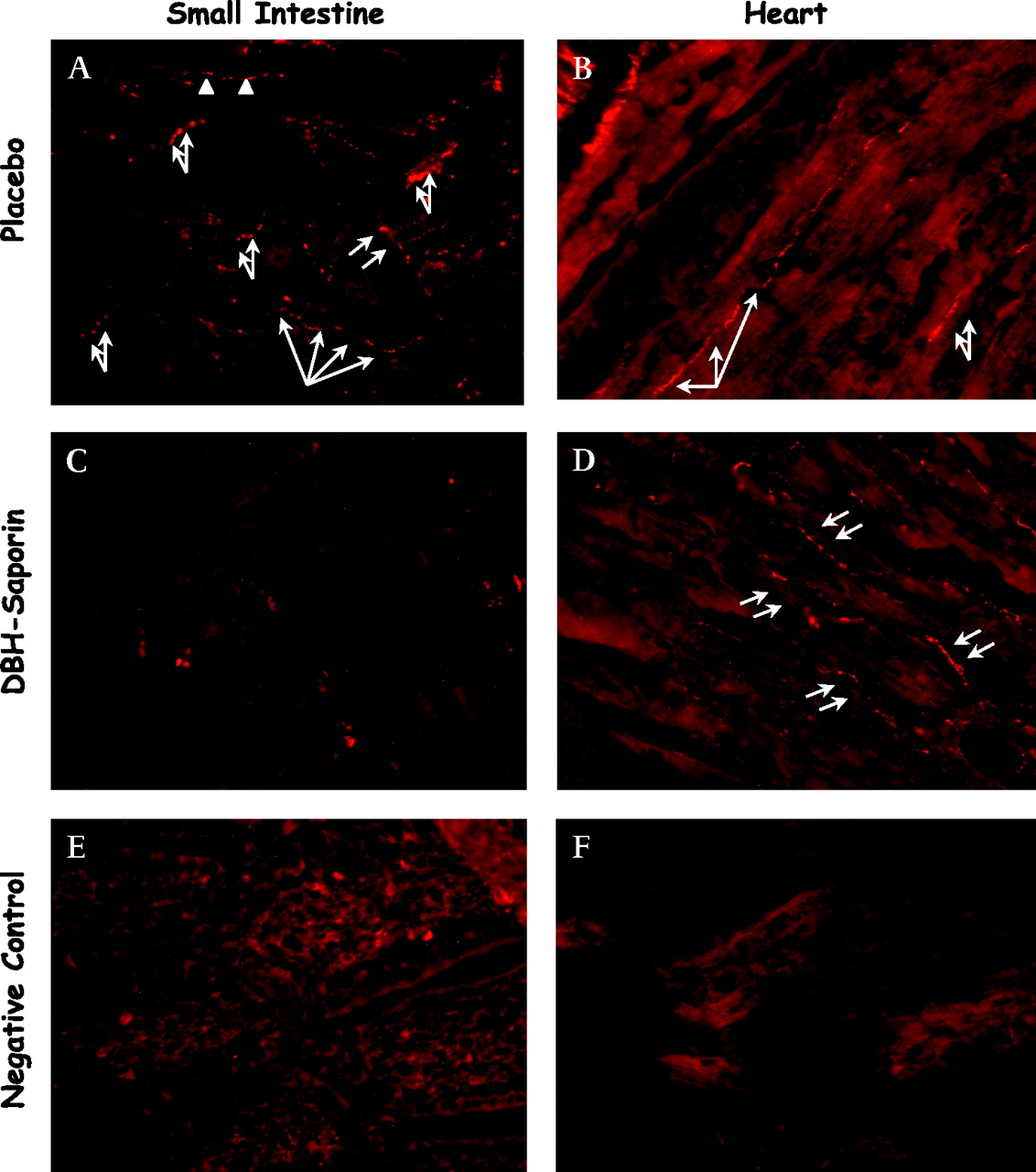
Denervation of splanchnic sympathetic nerves was verified by immunohistochemistry (figure 1). Representative images clearly indicate almost complete abrogation of sympathetic innervation in the small intestine and spleen (not shown). In contrast, sympathetic innervation of the heart remained unchanged, demonstrating the specificity of sympathectomy for the splanchnic area. Quantitative evaluation of the numbers of TH+ sympathetic single nerve fibres per square millimetre showed a marked decrease (>80% reduction) of TH+ sympathetic nerve fibres from the intestine in DBH immunotoxin-treated animals compared with saline-treated healthy rats (1.4±0.4 vs 8.3±2.3, p<0.001). In contrast, quantitative evaluation of TH+ nerve fibres per square millimetre in the heart showed no difference between animals with and without SE (7.0±1.8 vs 7.2±1.8).

Representative immunohistochemistry of sympathetic tyrosine hydroxylase nerve positive fibres of small intestine and heart in cirrhotic rats. (A) Small intestine without prior splanchnic sympathectomy; (B) heart without prior splanchnic sympathectomy; (C) small intestine with prior splanchnic sympathectomy; (D) heart with prior splanchnic sympathectomy; (E) negative control for small intestine; (F) negative control for heart. Magnification ×40. Sympathetic nerve fibres were completely abrogated after chemical sympathectomy in comparison with NaCl-treated animals. Arrows indicate sympathetic nerve fibres.
Translocation and spread of E coli
Protocol I
None of the cirrhotic ascitic rats presented with SBP or positive culture of peripheral blood, lung, liver or spleen. This is in accordance with previous investigations in the same animal model.27 40 Five of 11 cirrhotic rats (45%) showed bacterial translocation to MLN whereas only 2/12 cirrhotic rats with prior splanchnic SE (16.6%) were found to have positive culture of MLN (p<0.05).
Protocol II
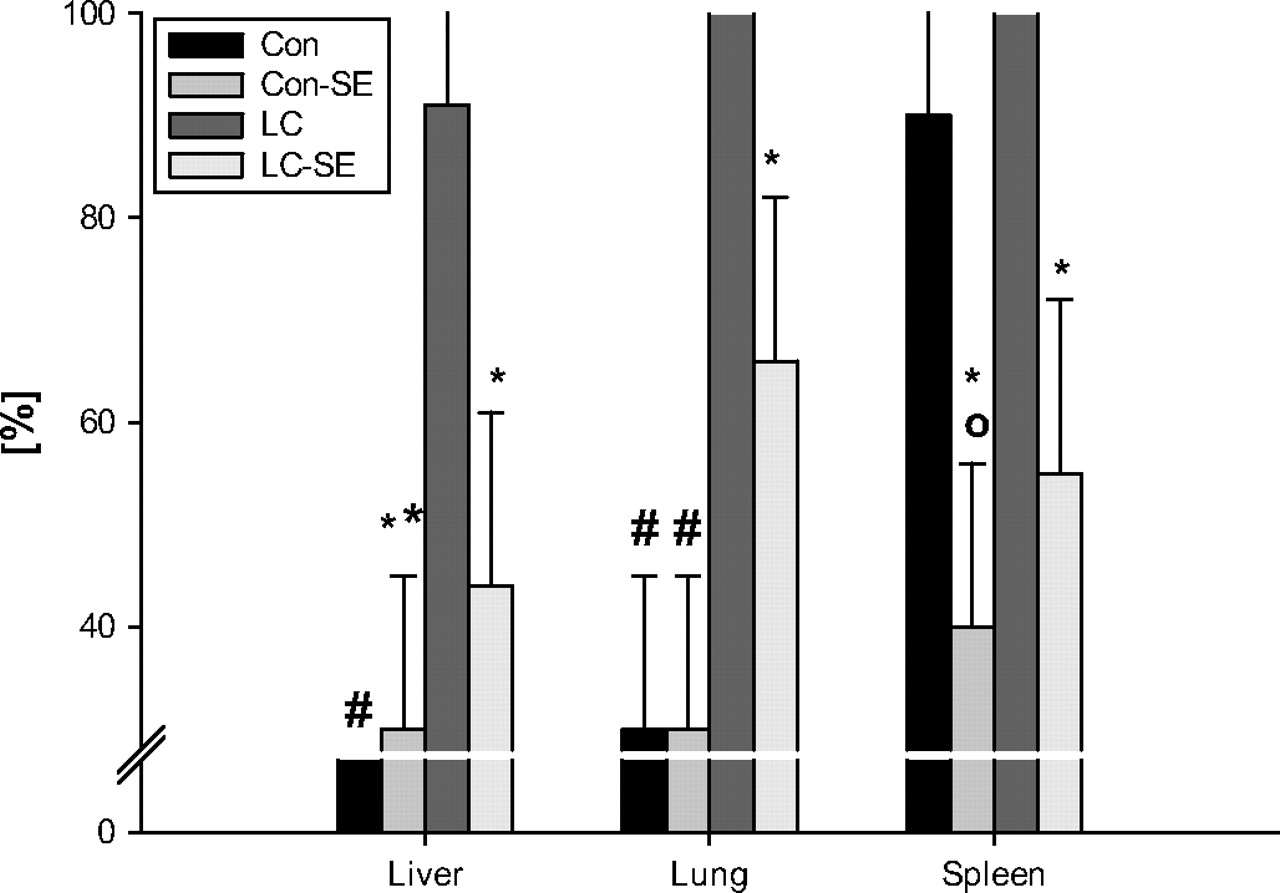
No cirrhotic or healthy control rats died during the observation period of 6 h after intraperitoneal E coli challenge. This is in accordance with the results obtained by Sanchez et al using a similar animal model.35 All cirrhotic rats with ascites showed translocation of injected E coli into MLN compared with only 20% of healthy control rats (table 2). Moreover, spread of E coli to the liver and lung occurred more frequently in cirrhotic ascitic rats than in control animals (figure 2). In addition, positive blood cultures were seen in cirrhotic rats but not in control rats (table 2). The bacterial tissue burden was markedly increased in the liver, spleen and lung of cirrhotic ascitic rats compared with control animals, underscoring the increase in the spread of E coli in these animals (figure 3).
Incidence of bacterial translocation of E coli and S aureus to mesenteric lymph nodes (MLN) and peripheral blood in control and cirrhotic rats (LC) with and without splanchnic sympathectomy (SE)

Incidence of Escherichia coli (intraperitoneal) translocation to liver, lung and spleen and effect of splanchnic sympathectomy (SE). *p<0.05, **p<0.01, #p<0.001 vs rats with cirrhosis (LC); °p<0.05 vs controls (Con).

Magnitude of bacterial spreading of Escherichia coli to liver, lung and spleen and effect of splanchnic sympathectomy (SE). (A) Organ-specific reduction in bacterial tissue burden by E coli after sympathectomy in cirrhotic rats. (B) Total in all organsyed. *p<0.05, **p<0.01, #p<0.001 vs rats with cirrhosis (LC). CFU, colony-forming units; Con, controls.
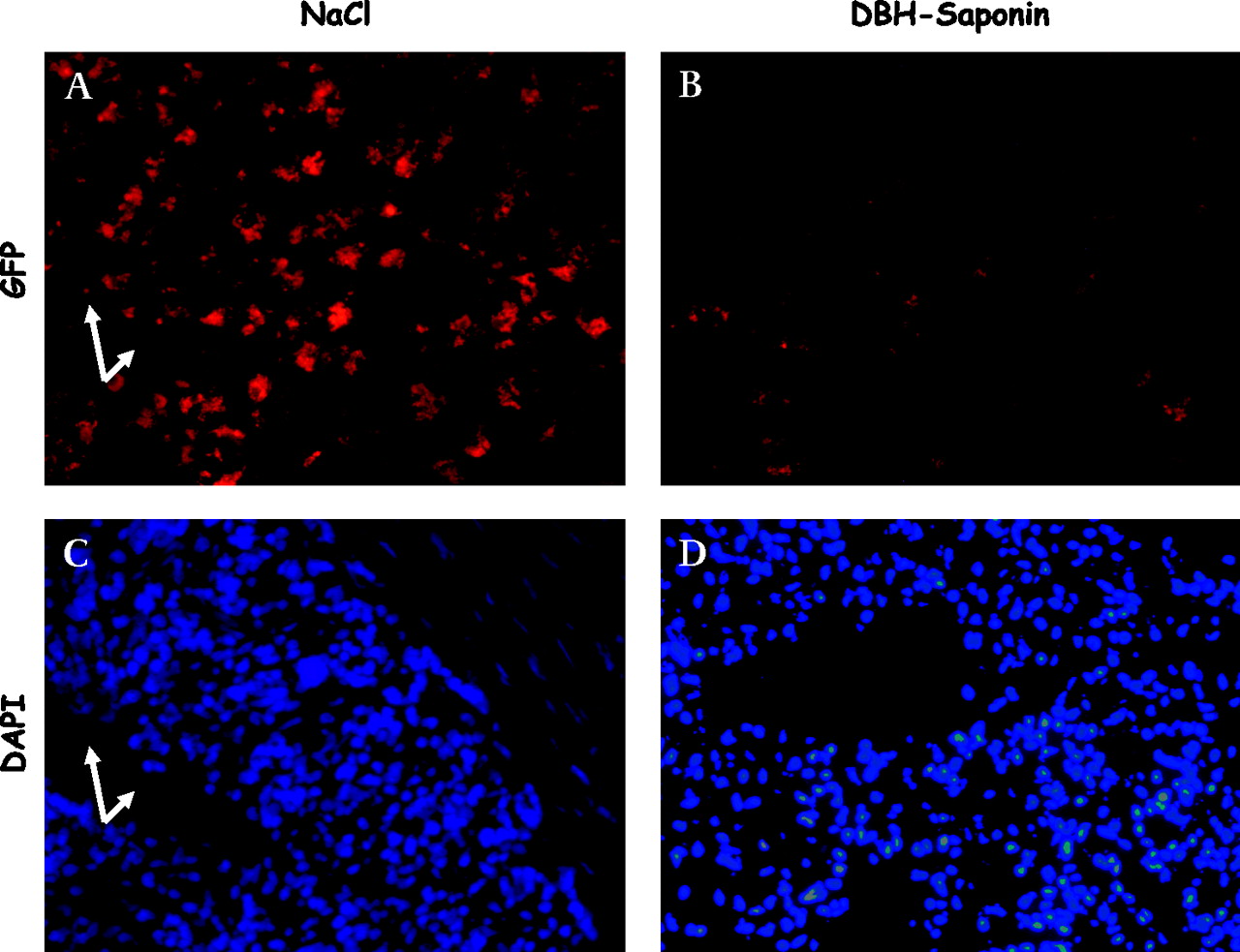
Ablation of the splanchnic sympathetic nervous system resulted in a marked reduction in the incidence of E coli translocation to MLN as well as to peripheral blood in cirrhotic ascitic rats (table 2). Only 1/9 cirrhotic rats with prior SE had bacteraemia compared with 7/11 cirrhotic rats without SE (p<0.05). Furthermore, the incidence of bacterial spreading to the liver, spleen and lung was significantly diminished (figure 2) and the bacterial tissue burden in the liver, spleen and lung were significantly reduced after SE in cirrhotic rats (figure 3). In fact, no significant difference in the incidence of bacterial translocation to MLN, liver, spleen, lung or blood or in tissue burden of bacteria was detectable between study groups after SE. In control rats the incidence and degree of bacterial translocation to MLN, liver and lung was not affected by splanchnic SE. However, the incidence of E coli translocation to the spleen was significantly ameliorated and the splenic tissue burden tended to be decreased (p=0.06) in control rats with prior SE compared with healthy controls. To visualise the influence of SE on the bacterial tissue burden, GFP-transfected E coli were used to infect control and cirrhotic rats, with and without splanchnic SE, and the spleens were removed 6 h later. Ablation of the splanchnic sympathetic nervous system caused a clear reduction in E coli-GFP in the spleen (figure 4).

Effect of splanchnic sympathectomy on splenic accumulation of green fluorescent protein (GFP)labelled Escherichia coli. (A, B) E coli-GFP in cirrhotic ascitic rats without prior splanchnic sympathectomy (NaCl treated; A) and with prior splanchnic sympathectomy (DBH-Saporin; B). Arrowheads show accumulation of GFP-labelled E coli. The high-power fields in panels C and D show tissue stained with 4′6′-diamidino-2-phenylindole hydrochloride (DAPI) identifying cell nuclei and splenic tissue structures. The cell-rich areas are T lymphocyte areas of the periarteriolar lymphatic sheet with the central artery in the middle (arrows).
Translocation and spreading of S aureus
Protocol III
Neither cirrhotic nor healthy control rats died during the observation period of 6 h after intraperitoneal S aureus challenge. All cirrhotic rats with ascites showed translocation of injected S aureus into MLN compared with only 56% of healthy control rats (table 2). In cirrhotic rats the incidence of spread of S aureus to the liver and spleen was similar to that in control rats but tended to be increased in the lung (data not shown). Moreover, the severity of spreading of S aureus to MLN, liver, spleen and lung tended to be increased in cirrhotic rats but the difference did not achieve statistically significant levels (data not shown). S aureus bacteraemia occurred more frequently in cirrhotic ascitic rats than in control animals (table 2). In cirrhotic ascitic rats but not in healthy controls, translocation of endogenous Gram-negative bacteria was observed to MLN, liver, spleen and lung (figure 5). Prior ablation of the splanchnic sympathetic nervous system was not associated with any significant change in the incidence or severity of S aureus translocation to MLN, liver, spleen, lung or blood in cirrhotic or healthy rats (data not shown). However, splanchnic SE did reduce the incidence of Gram-negative and thus endogenous bacterial translocation from the gut to MLN, liver, spleen and lung in cirrhotic rats but this did not reach statistical significance (figure 5).
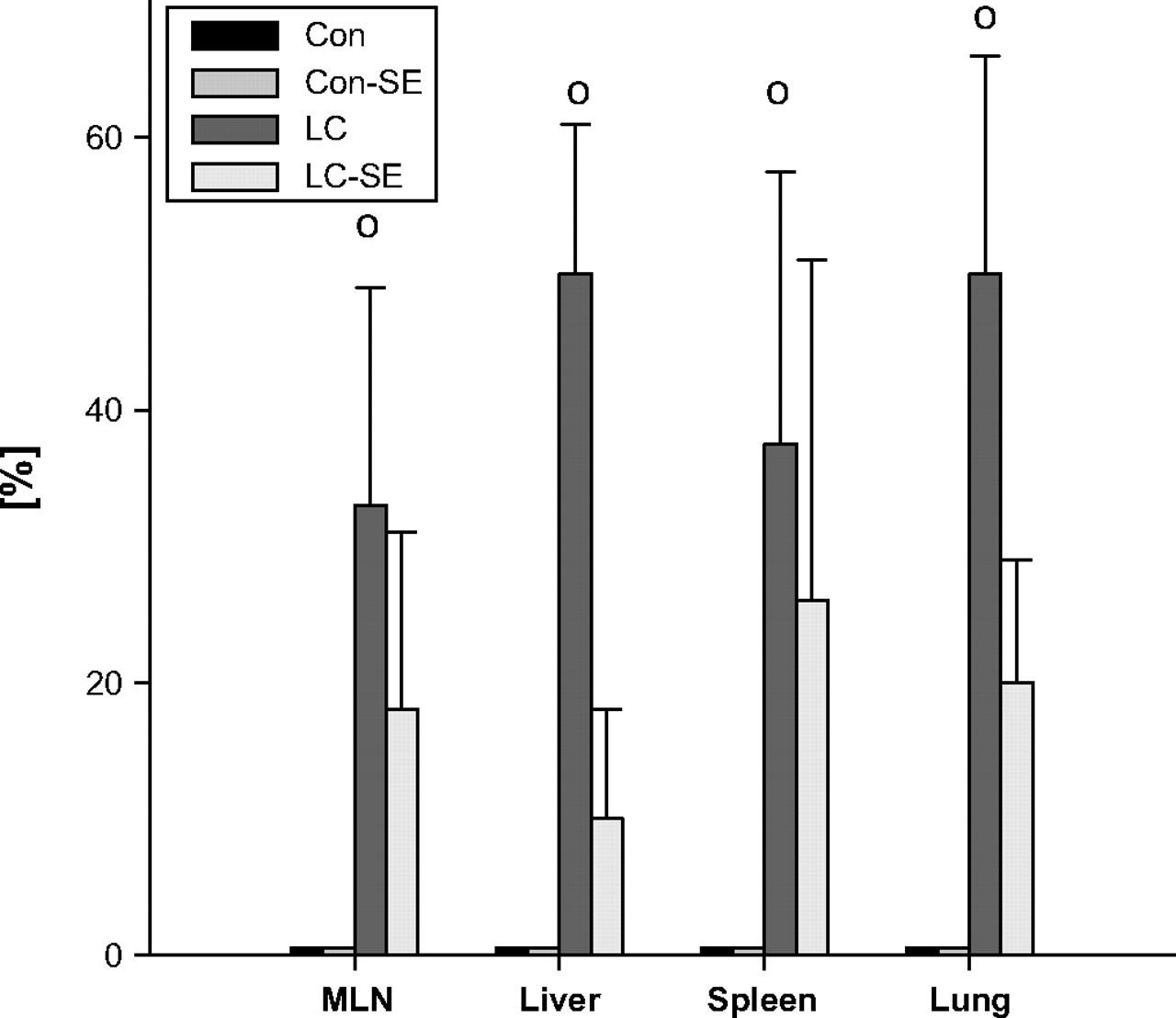
Incidence of endogenous Gram-negative translocation to mesenteric lymph nodes (MLN), liver, lung and spleen in cirrhotic and healthy control rats after intraperitoneal Staphylococcus aureus challenge and its dependency on splanchnic sympathectomy. οp<0.05 versus control. Con, controls; LC, rats with cirrhosis; SE, splanchnic sympathectomy.
Influx of PMNL into the peritoneal cavity and cytokine profile after E coli and S aureus intraperitoneal challenge
E coli resulted in a significantly greater influx of PMNL into the peritoneal cavity than S aureus (table 3). Ablation of the splanchnic sympathetic nervous system increased the influx of monocytes and neutrophils into the peritoneal cavity in cirrhotic ascitic rats in response to intraperitoneal E coli but not in response to S aureus. No change in the numbers of lymphocytes was observed in peritoneal fluid after E coli or S aureus challenge by prior sympathectomy in cirrhotic ascitic rats.
Numbers of neutrophils, monocytes and lymphocytes in the peritoneal cavity of cirrhotic rats with ascites with and without splanchnic sympathectomy (SE) after intraperitoneal E coli or S aureus challenge
In vitro PMNL phagocytic activity of E coli
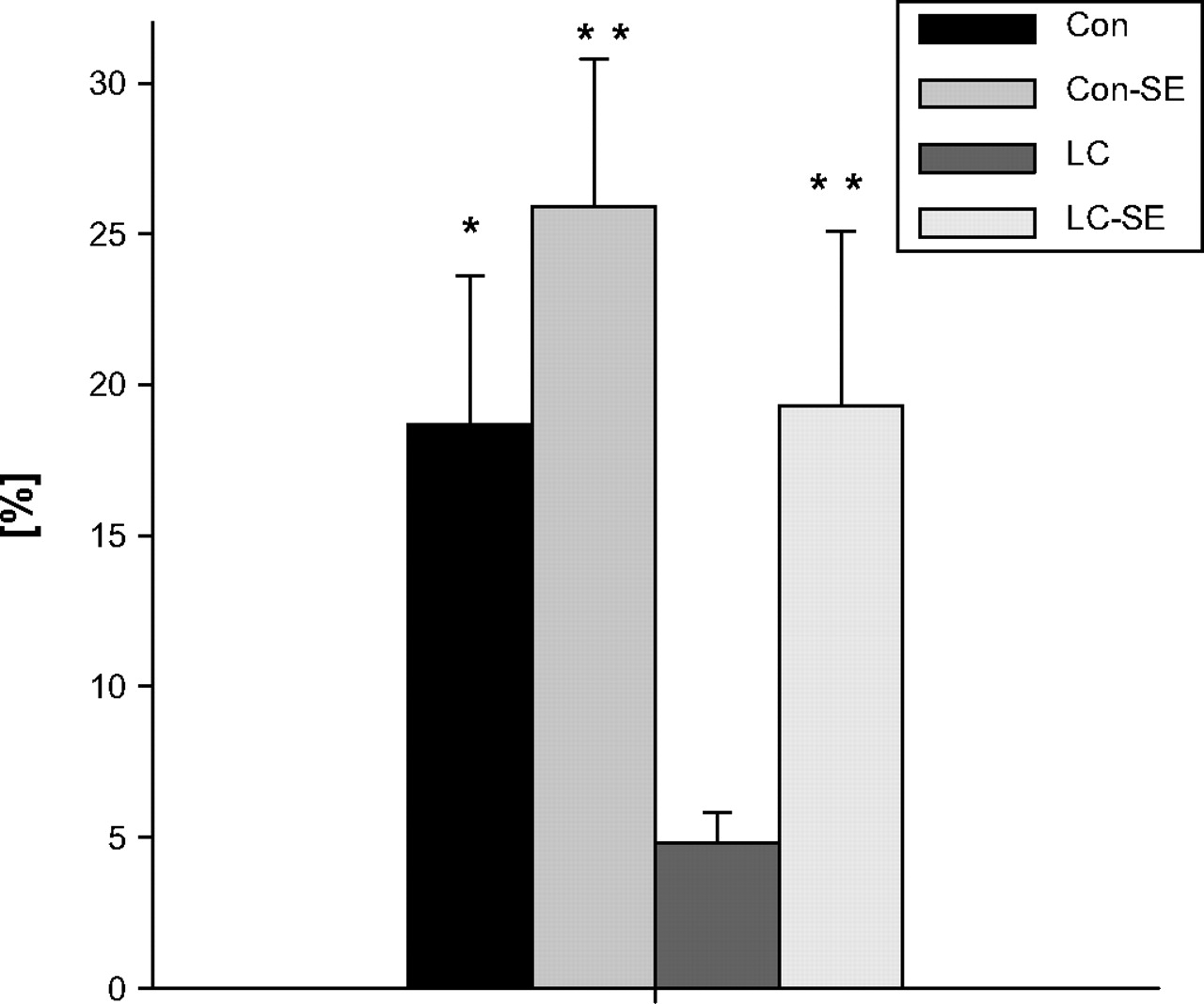
Autofluorescence of PMNL was <101 fluorescence intensity. No difference in mean fluorescence intensity >101 at baseline (4°C) given by intracellular FITC-labelled E coli was observed between the study groups. After an incubation period of 10 min at 37°C, comparison of mean fluorescence activity (FL1) in PMNL between the study groups showed no significant difference (data not shown). The percentage of PMNL contributing to FL1 as determined by M(a) was found to be significantly decreased in cirrhotic animals compared with healthy control rats. This so-called readiness of peritoneal PMNL to phagocytose was significantly increased in cirrhotic rats by prior SE and no more different from healthy control rats (cirrhotic rats with prior SE 27.6±5.7%, control rats 27.9±5.2%, control rats with prior SE 38.9±5.6% vs 4.8±0.7% in cirrhotic rats, all p<0.001). Likewise, the fraction of active PMNL tended to be increased in control rats after SE but this difference did not reach statistical significance. The phagocytic index of peritoneal PMNL was markedly attenuated in cirrhotic ascitic rats compared with healthy controls (figure 6). Prior ablation of the splanchnic sympathetic nervous system led to a significantly enhanced phagocytic index in cirrhotic ascitic rats which was no more different from control animals with or without prior SE.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In vitro phagocytosis index of peritoneal polymorphonucelar leucocytes (PMNL) for Escherichia coli. *p<0.05, **p<0.01 vs rats with cirrhosis. Con, controls; LC, rats with cirrhosis; SE, splanchnic sympathectomy.
Discussion
Splanchnic SE prevents spontaneous bacterial translocation to MLN in cirrhotic rats with ascites. Although not studied, this can be attributed to proposed effects of SE resulting in acceleration of intestinal transit time,41 42 prevention of Gram-negative intestinal bacterial growth43 or improvement in gastrointestinal permeability.44 However, by applying an experimental model of bacterial peritonitis, splanchnic SE decreased the incidence and severity of systemic spreading of E coli. Additional effects mediated by SE are therefore necessary to explain this finding.
The sympathetic nervous system, mainly via norepinephrine, exerts potent immunosuppressive actions including inhibition of chemotaxis/migration of monocytes and neutrophils19 45 and blockade of phagocytosis of bacteria20 21; these activities are particularly essential for defence against Gram-negative bacteria. Prior splanchnic SE resulted in an increased influx of PMNL into the peritoneal cavity of cirrhotic ascitic rats, confirming previous reports of the effect of SE on leucocyte influx into the peritoneal cavity.19 22 Stimulation of neutrophilic adrenergic receptors has been shown to decrease neutrophil adhesion and expression of CD11b resulting in the prevention of PMNL recruitment and function.46 Splanchnic SE resulted in increased phagocytic activity of PMNL in cirrhotic ascitic rats which was due mainly to an improvement in the readiness of PMNL to phagocytose E coli resulting in a higher number of PMNL involved in the phagocytic process. The SE-induced decrease in organic bacterial numbers and incidence of bacteraemia after intraperitoneal E coli challenge may therefore be due to increased destruction of E coli in the peritoneum before bacterial entry into the circulation, mediated by an increase in the influx and phagocytic activity of PMNL.
These results are in accordance with recent studies in mice.22 47 48 Straub et al reported that ablation of the sympathetic nervous system decreased the bacterial tissue burden in the spleen48 and also in the liver and lung of experimental animals after intraperitoneal challenge with E coli.22 Moreover, chemical SE reduced the splenic bacterial load after intraperitoneal injection of Listeria monocytogenes.47 In addition, treatment with adrenergic agonists increases mortality during Gram-negative bacterial infections.49 Similarly, activation of the sympathetic nervous system by experimental stress led to an increased tissue burden of Gram-negative bacteria.50 This effect was markedly inhibited by propranolol, indicating that β-adrenergic signalling mediates the lack of resistance to Gram-negative bacteria. Propranolol has also been used in experimental cirrhosis and was found to lower the rate of bacterial translocation.41
Interestingly, intraperitoneal challenge with S aureus most likely causing severe sepsis resulted in endogenous bacterial translocation of Gram-negative bacteria from the gut to the MLN, liver, spleen and lung in cirrhotic rats but not in healthy controls. This demonstrates a greater susceptibility to gut-derived Gram-negative infections in cirrhosis via bacterial translocation. In fact, this can easily be explained by the presence of intestinal overgrowth of Gram-negative bacteria, increased intestinal permeability and impaired host defence in these animals compared with healthy controls. Likewise, the increased systemic spread of E coli and S aureus after intraperitoneal application in liver cirrhosis can be explained by the well-known depression of the hepatic reticuloendothelial system present in advanced liver disease. The failure of the liver in ‘filtering’ these bacteria allows passage of bacteria from the portal venous system to the systemic circulation. Thus, the risk of acquiring bacteraemia and SBP in cirrhosis is directly related to the degree of dysfunction of the reticuloendothelial system.51 However, marked differences in host defence dependent on the invading organism are apparent.52 53
Splanchnic SE had no significant effect on systemic spreading of S aureus from the peritoneal cavity to the systemic circulation. Moreover, the influx of PMNL into the peritoneal cavity in response to S aureus was not altered by SE. This may be due to the fact that S aureus expresses a chemotaxis inhibitory protein that specifically impairs the response of neutrophils and monocytes to formylated peptides and C5a.54 Furthermore, Gram-positive infections induce mainly interleukin 4 secretion, release of corticosteroids and influx of lymphocytes, shaping the immune response.22 However, the influence of the sympathetic nervous system on these effector systems is low and rather stimulatory.22 Of note, the number of neutrophils recruited to the peritoneal cavity was significantly lower in response to S aureus than to E coli in cirrhotic rats. This is in accordance with reports on Gram-positive SBP in patients with cirrhosis presenting with PMN counts below the threshold of 250/mm3.55 Moreover, bactDNA from Gram-negative bacteria in ascitic fluid is associated with a higher ascitic PMN count than the presence of bactDNA from Gram-positive bacteria,56 underscoring the differences in stimulatory capacity for PMN dependent on the strain of bacteria under consideration.
Our results shed new light on previous observations that the well-known decrease in mortality induced by non-selective β blockade may be at least in part due to its preventive effect on the incidence of infectious complications.57 E coli is one of the most frequently isolated bacteria causing SBP, particularly when community-acquired.10 58 The use of β blockers in advanced liver cirrhosis has recently been reported to lower the probability of community-acquired SBP,59 and a recent meta-analysis concluded that this protective effect is even independent of the haemodynamic response achieved.60 The current data therefore provide evidence that these beneficial effects of β blockers in patients with cirrhosis may be mediated by a decrease in bacterial translocation and spreading of Gram-negative bacteria induced by modulation of the host defence.
In summary, this study suggests that sympathetic nervous system hyperactivity in the splanchnic circulation increases susceptibility to bacterial translocation and infections with E coli but not S aureus. This detrimental effect seems to be due to suppression of the innate immune response to Gram-negative bacteria mediated by the sympathetic nervous system, thus altering the ability of the host to eliminate the pathogen.
Acknowledgments
The authors gratefully acknowledge the excellent technical assistance of Angelika Gräber and Anja Szabo.
References
Footnotes
Funding This work was supported by a grant from Deutsche Forschungsgemeinschaft DFG (Wi 1502/5-2) to RW.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Digest