Article Text

Abstract
It is increasingly perceived that gut host–microbial interactions are important elements in the pathogenesis of functional gastrointestinal disorders (FGID). The most convincing evidence to date is the finding that functional dyspepsia and irritable bowel syndrome (IBS) may develop in predisposed individuals following a bout of infectious gastroenteritis. There has been a great deal of interest in the potential clinical and therapeutic implications of small intestinal bacterial overgrowth in IBS. However, this theory has generated much debate because the evidence is largely based on breath tests which have not been validated. The introduction of culture-independent molecular techniques provides a major advancement in our understanding of the microbial community in FGID. Results from 16S rRNA-based microbiota profiling approaches demonstrate both quantitative and qualitative changes of mucosal and faecal gut microbiota, particularly in IBS. Investigators are also starting to measure host–microbial interactions in IBS. The current working hypothesis is that abnormal microbiota activate mucosal innate immune responses which increase epithelial permeability, activate nociceptive sensory pathways and dysregulate the enteric nervous system. While we await important insights in this field, the microbiota is already a therapeutic target. Existing controlled trials of dietary manipulation, prebiotics, probiotics, synbiotics and non-absorbable antibiotics are promising, although most are limited by suboptimal design and small sample size. In this article, the authors provide a critical review of current hypotheses regarding the pathogenetic involvement of microbiota in FGID and evaluate the results of microbiota-directed interventions. The authors also provide clinical guidance on modulation of gut microbiota in IBS.
- Functional gastrointestinal disorders
- microbiota
- irritable bowel syndrome
- small intestinal bacterial overgrowth
- breath tests
- probiotics
- prebiotics
- synbiotics
- antibiotics
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/2.0/ and http://creativecommons.org/licenses/by-nc/2.0/legalcode.
Statistics from Altmetric.com
- Functional gastrointestinal disorders
- microbiota
- irritable bowel syndrome
- small intestinal bacterial overgrowth
- breath tests
- probiotics
- prebiotics
- synbiotics
- antibiotics
Introduction
Functional gastrointestinal disorders (FGIDs) are defined by symptom-based diagnostic criteria that combine chronic or recurrent symptoms attributable to the GI tract in the absence of other pathologically-based disorders.1 The FGIDs are classified into six major categories for adults: oesophageal, gastroduodenal, bowel, functional abdominal pain syndrome, biliary and anorectal. Of these, the functional bowel disorders (FBD) constitute one of the most common reasons for seeking healthcare,2 and they are associated with poor health-related quality of life3–5 and substantial costs to society.6–9 The pathophysiological mechanisms underlying these disorders are incompletely known, but abnormal gastrointestinal (GI) motility, visceral hypersensitivity, altered brain–gut function, low-grade inflammation, psychosocial disturbance and intestinal microbes may contribute.10–12
The human body is inhabited by a complex community of microbes, collectively referred to as microbiota.13 It is estimated that the human microbiota contains 1014 cells, which outnumber the human cells in our bodies by a factor of ten.14 A vast majority of these are found in the GI tract, with a continuum from 101–103 bacteria per gram of content in the stomach and duodenum to 1011–1012 cells per gram in the colon.15 Moreover, the microbial composition differs between these sites,16 and there are also significant differences between the microbiota present in the gut lumen and the microbiota attached to and embedded in the mucus layer of the GI tract.17 The microbiota is taxonomically classified via the traditional biological nomenclature (phylum—class—order—family—genus—species) and currently more than 50 bacterial phyla have been described, of which 10 inhabit the colon and three predominate: the Firmicutes, Bacteroidetes and the Actinobacteria; other sites display a different microbial composition.18 ,19 A challenge for researchers and clinicians is that most of the microbial diversity in the human GI tract is not currently represented by available cultured species,20 but during recent years, the use of culture-independent techniques to study the gut microbiota has increased the understanding of the role of gut microbiota in health and disease.14
Several lines of evidence indicate that bacteria may be involved in the pathogenesis and pathophysiology of FBD, through the metabolic capacity of the luminal microbiota, and the potential of the mucosa-associated microbiota to influence the host via immune–microbial interactions.21 For instance, many subjects with irritable bowel syndrome (IBS) report symptom onset following an enteric infection.22 There are also studies reporting positive effects of treatments directed at gut microbiota in patients with FBD.23 ,24 Moreover, small intestinal bacterial overgrowth (SIBO)25 and altered intestinal microbiota26 are implicated in subgroups of FBD patients. However, the clinical relevance of these findings remains unclear and, therefore, we sought to critically review the existing literature on the role of intestinal microbiota in FBD, focusing predominantly on IBS, and to provide recommendations for how to implement the current knowledge into clinical practice and to guide future research.
This manuscript is a synthesis of the endeavour of the Rome Foundation Committee Report. More indepth description of the work produced by this team is provided as online supplementary material.
Current knowledge of the microbiota
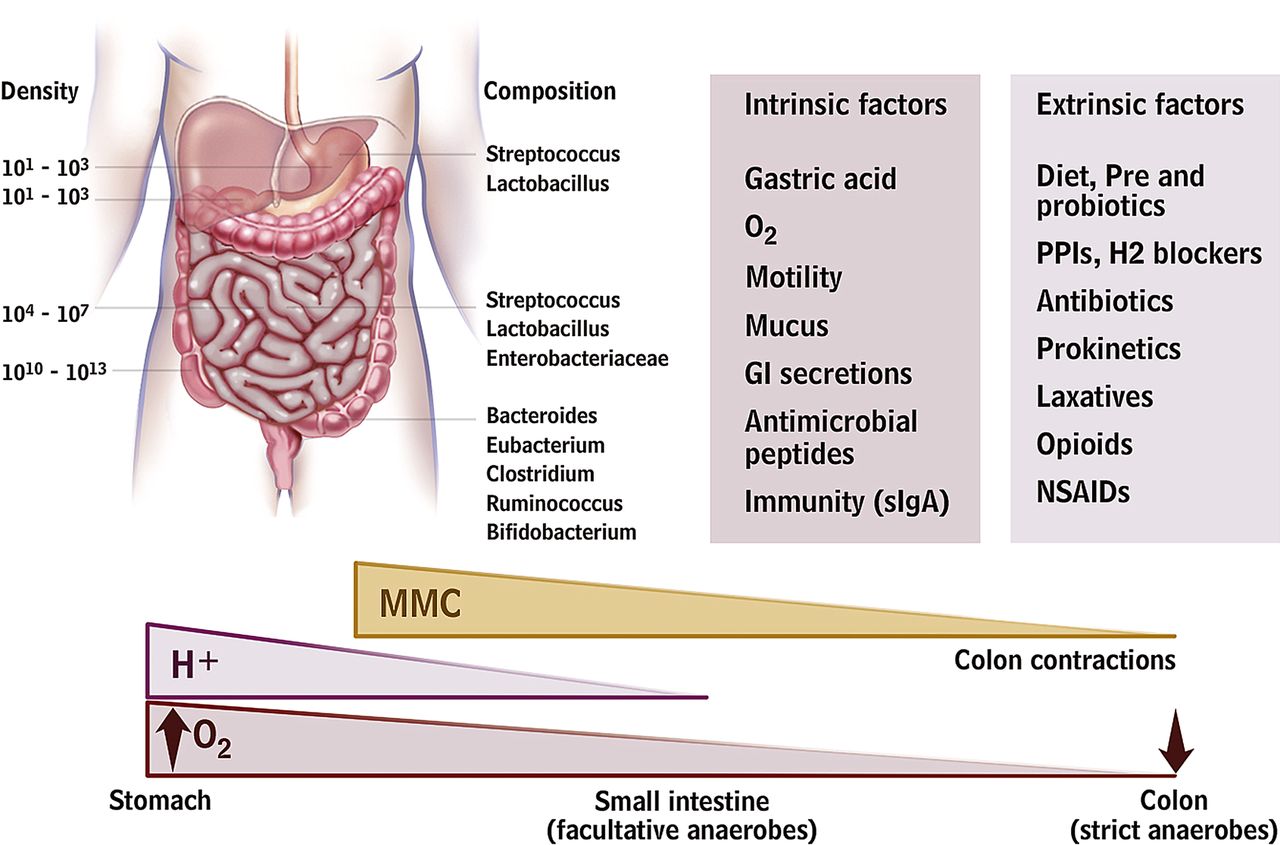
A relationship, often termed symbiosis, has developed between the host and the intestinal microbiota over millions of years. Host genetic and immune as well as environmental factors influence intestinal microbiota composition which in turn shape host immunity and physiology within and beyond the gut (figure 1). Recent human studies demonstrate a hitherto unimagined complexity of the human gut microbiota with hundreds of phylotypes, of which 80% remain uncultured.19 Of the 10 bacterial phyla detected in the gut the Firmicutes, Bacteroidetes and Actinobacteria predominate, of which the Firmicutes is the most dominant and diverse phylum in the GI tract. Facultative anaerobes account for <0.1% of the total bacteria detected in faecal samples. A recent paper suggested that the human GI tract microbiota can be divided into three robust clusters called enterotypes formed by groups of species that jointly contribute to their respective preferred community composition.27 Remarkably, these enterotypes do not vary by patient characteristics, such as nation, gender, age or body mass index, although these findings are based on relatively small numbers of subjects. While most studies used faecal material, this does differ somewhat from the bacteria adherent to the mucosa, which are likely to interact most strongly with the host.28

Gut microbiota and the intrinsic and extrinsic factors that can affect its distribution and composition. A number of host mechanisms participate in gut microbiota modulation, including gastric acid secretion, fluid, anticommensal sIgA and antimicrobial peptide production, and gastrointestinal (GI) motility. Drugs that block acid secretion and affect GI motility can indirectly alter the microbiota. Antibiotics, depending on spectrum and dosage, will directly affect microbiota composition. Dietary modifications, including probiotic and fibre supplements, will also affect microbiota composition. MMC, migrating motor complexes; H+ hydrogen ions; O2, partial oxygen tension; sIgA, secretory immunoglobulin A; PPI, proton pump inhibitor; NSAID, non-steroidal anti-inflammatory drug.
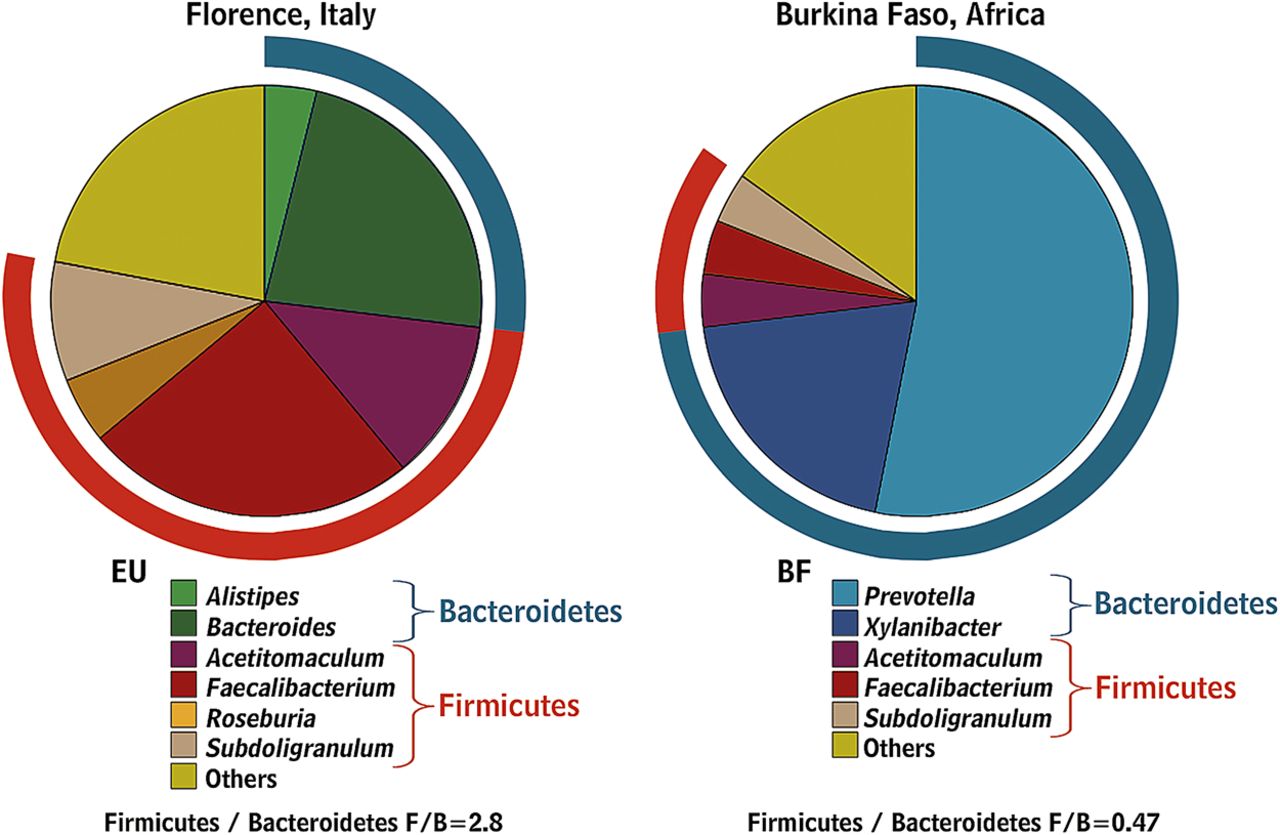
Babies are born with sterile intestines but are rapidly colonised by bacteria from their immediate environment, most importantly their mother's vagina and gut.29 Early colonisers of the neonatal gut are mainly aerobes (such as staphylococci, streptococci and enterobacteria), while late colonisers are strict anaerobes (such as eubacteria and clostridia) as the total microbiota become more complex, more stable and converge to a common pattern.30 ,31 The microbiota continue to evolve until adulthood with a gradual increase in Bacteroides spp., a decline in Lactobacillus spp. after the age of five and a decline in Bifidobacterium spp. in late teenage.32 Changes also occur in extreme old age when Bacteroides spp. decrease while Enterococcus spp. and Escherichia coli increase.33 ,34 Industrialisation has changed both our diet and microbiota as evidenced by comparing the faecal microbiota of African rural children with a polysaccharide-rich diet with Italian city children on a high fat, high protein diet. African children have a significant enrichment in Bacteroidetes, especially Prevotella and Xylanibacter genera known to contain genes for xylan hydrolysis35 (figure 2). Whole grain cereals,36 resistant starch37 ,38 and low residue diets profoundly alter the microbiota.39 Although there is evidence indicating that obese individuals have an increase in Firmicutes and a decrease in Bacteroidetes (a difference likely related in part to different diets40), other studies failed to support these observations.41 ,42 Many dietary prebiotics including oligofructose,43 lactulose,44 ,45 lupin kernel,46 inulin-containing juices47 and arabinoxylan-oligosaccharides48 significantly alter human faecal microbiota. The concept of poorly absorbed but fermentable oligo-, di- and mono-saccharides and polyols (FODMAPs) includes many substances which are substrates for bacterial metabolism and may therefore alter the microbiota but this has as yet not been studied.
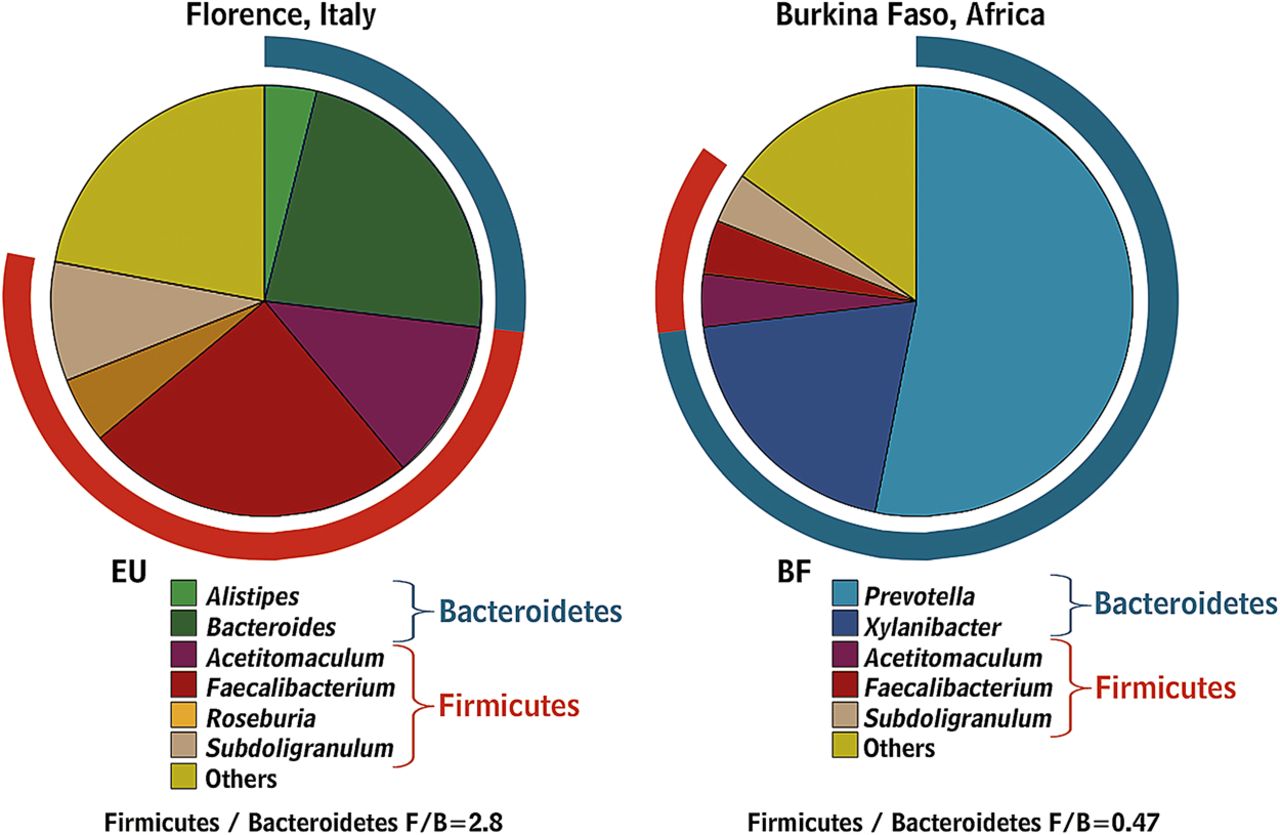
Gut microbiota composition in African children living in rural areas with a polysaccharide-rich diet when compared with Italian city children.35 (Reprinted with permission from Proc Natl Acad Sci USA).
Most high fibre diets alter the microbiota and accelerate transit. Accelerating transit using senna increased the production of short chain fatty acids (SCFAs) but reduced faecal methanogens, the opposite to the effect of loperamide.49 Accelerating transit with cisapride also increases production of SCFAs, particularly propionic and butyric acids.50 Acetate, which predominates in the colonic contents, is largely inhibitory. In contrast, propionate and butyrate stimulate motility, activate propulsive ileal motor patterns in humans51 and ensure that bacteria are propelled from the ileum to the colon. The normal microbiota also strongly influence the mucosal immune system52 ,53 which is underdeveloped in germ-free animals, who have reduced T cells, immunoglobulin A producing B cells and intraepithelial T cells.52 ,54–56 Twin studies suggest that the host genotype influences the gut microbiota, although results remain conflicting because of the inability to control for shared environmental factors.40 ,57 One of the most important genetic effects is mediated via the innate immune response. Thus, mice lacking the bacterial sensing receptor nucleotide-binding oligomerisation domain-containing protein-2 showed significantly more Bacteroidetes as well as Firmicutes compared with wild-type mice.58
Modulation of the microbiota induces visceral hypersensitivity in mice, which is reduced by Lactobacillus paracasei NCC2461 secreted products.59 Lactobacillus acidophilus NCFM and Lactobacillus paracasei NCC2461 also modulate visceral pain perception in rodents.60 ,61 Transient perturbation of the microbiota with antimicrobials alters brain-derived neurotrophic factor expression, exploratory behaviour and colonisation of germ-free mice suggesting that intestinal microbiota impact is not limited to the gut and the immune system, but may involve the central nervous system.62 (Note: this last sentence appears run-on but I can't quite decipher how to fix it.)
Approaches to the study of microbiota
Approaches to the study of microbiota and relative advantages/pitfalls are reported in box 1. Culture-based studies reveal that the gut microbiota is a highly complex community (box 1).63 Although culturing remains valuable for identifying functional groups and for selective enumeration (eg, of pathogens), new culture-independent approaches provide more powerful and convenient methodologies for monitoring changes in the GI tract community (table 1). Information on the diversity of microbes that colonise the gut has expanded rapidly over the past 15 years, based largely on the analysis of the small subunit ribosomal RNA (16S rRNA for Bacteria and Archaea, 18S rRNA for Eukaryotes) gene sequences that can be obtained by direct amplification from nucleic acids extracted from gut or stool samples.64 This information provides the basis for a range of complimentary techniques for enumerating gut bacteria, including fingerprinting methods such as denaturing gradient gel electrophoresis65 and targeted methods such as fluorescent in situ hybridisation and quantitative PCR. The arrival of new high-throughput sequencing approaches and 16S rRNA-based microarraying has further accelerated the supply of data by allowing amplified 16S rRNA sequences to be analysed indepth without the need for ‘classical’ cloning and sequencing methods.66 ,67 Although culturing may bias against bacteria that are hard to grow in the laboratory, PCR amplification biases against certain groups of gut bacteria. For example, bifidobacterial 16S rRNA sequences are often under-represented among amplified products, although more reliably enumerated by 16S rRNA-targeted fluorescent in situ hybridisation detection or quantitative PCR.37 While most molecular enumeration methods target 16S rRNA, some are based on more functionally relevant genes, for example, involved in methanogenesis or butyrate synthesis.
Box 1 Approaches to the study of intestinal microbiota
-
Breath tests are not validated to accurately detect small intestinal bacterial overgrowth.
-
Rapid molecular approaches have largely replaced cultural approaches for enumeration of the dominant gastrointestinal (GI) tract microbiota.
-
Cultural microbiology remains crucial for investigating microbial diversity and for the selective isolation of representatives of key functional groups, including pathogens.
-
Culture-independent approaches to study the GI tract microbiota can answer the questions:
-
Which microbes are present in the GI tract? (16S rRNA gene-based approaches)
-
What microbial genes are present in the GI tract? (metagenomics)
-
What are GI tract microbes doing? (metatranscriptomics, metaproteomics, metabonomics/metabolomics).
-
-
The possibilities of using high-throughput approaches and their depth of analysis are increasing rapidly, but it is important they are applied with careful reference to well-defined scientific questions.
Main features of culture-independent detection methods of gut microbiota
High-throughput DNA sequencing provides completely new possibilities for ‘-omics’-based analyses of the gut microbiota.19 Draft genomes of cultured gut bacteria can now be produced rapidly and at little cost.68 In addition, these methods can be applied to DNA recovered from gut or stool samples, and the analysis of the resulting complex mixture of sequences is referred to as metagenomics.69 ,70 The ability to analyse multiple gene sequences from large numbers of samples, complemented with functional screening and characterisation of randomly cloned DNA fragments from the GI tract, is currently being exploited to uncover changes in disease states including in inflammatory bowel disease (IBD). A related technology, metatranscriptomics, uses high-throughput sequencing or microarray analysis to examine RNA expressed in GI tract samples, thus focusing on bacteria that are transcriptionally active. Another potentially powerful tool, metaproteomics, employs protein separation and sequencing techniques to describe the major proteins present in gut or stool samples.71 ,72 These ‘meta-omics’ approaches rely in primary sequencing and annotation data.73 ,74 Thus, they rely heavily on the availability of genome sequences and functional information from cultured reference bacteria, which means there are considerable benefits from combining different approaches to gut microbiota analysis. A final ‘omics’ approach, metabonomics, is not linked directly to genetic information of the microbes, but examines the metabolite profiles resulting from total microbial activity in the gut. Since many of these metabolites exert biological effects (some positive, some negative) on the host, such analysis can provide a direct measure of the consequences of microbial activity in the gut, although excluding cell-mediated effects and direct identification to a target microbial species.
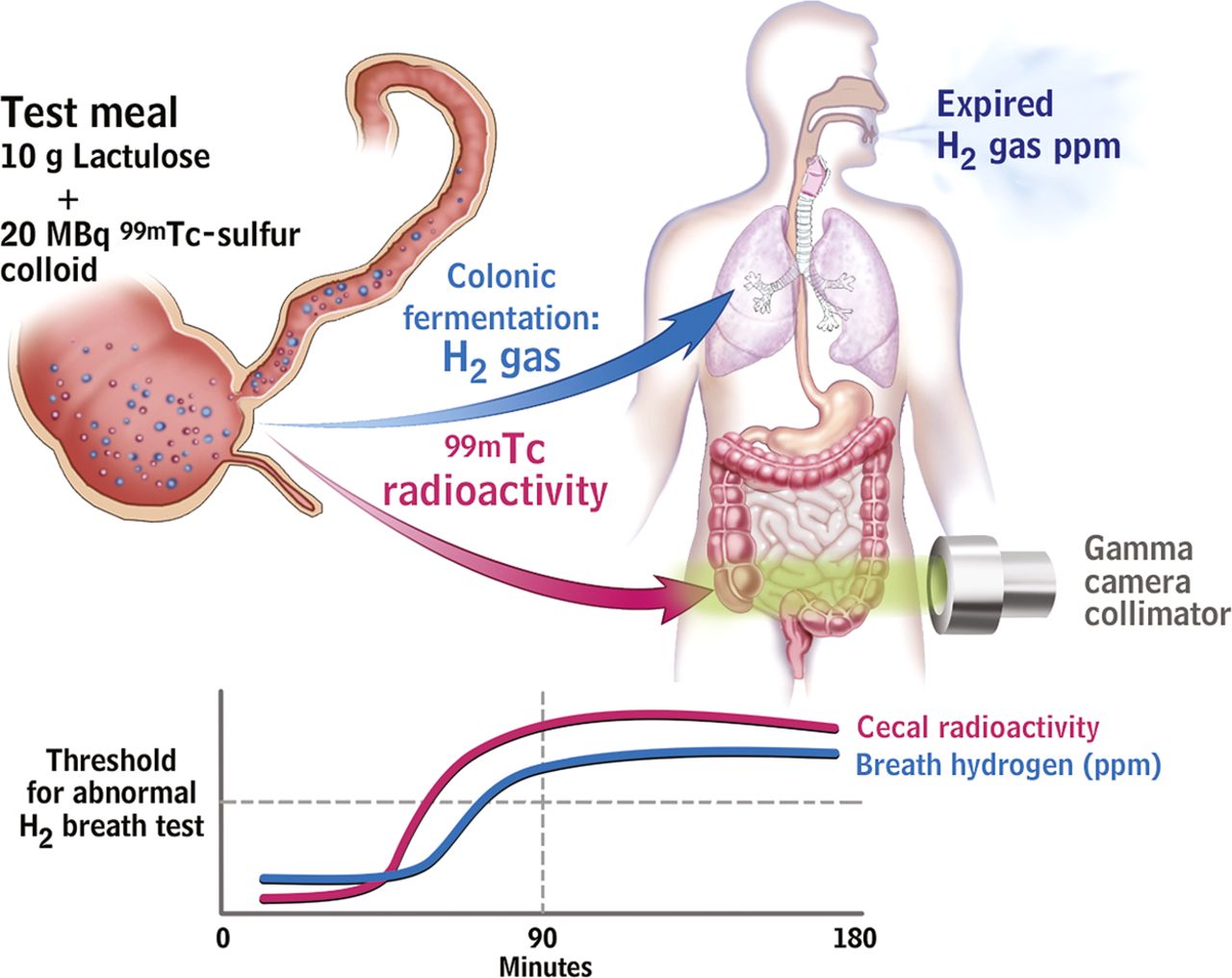
Breath testing has been used to detect SIBO in IBS patients by non-invasively detecting hydrogen producing bacteria or methane producing archaea within the gut lumen. The breath test is based on the concept that hydrogen gases are produced by colonic bacterial fermentation in response to ingestion of a test sugar. They rapidly diffuse into the blood, are excreted by breath, and can be collected and quantified.75 If SIBO exists, the timing of this fermentation would be altered but the criteria for abnormal tests lack validity (figure 3).

The lactulose hydrogen breath test (LHBT) predominantly measures small intestinal transit rather than small intestinal bacterial overgrowth (SIBO) in irritable bowel syndrome (IBS) patients. Upper schematic shows ingestion of test meal with subsequent serial measurement of both H2 gas, resulting from fermentation of the lactulose by intestinal bacteria, and Tc99 scanning in the caecum. This latter measurement detects when the test meal has reached the caecum. The stylised drawing below shows a representative result from an IBS patient with serial measurements over time. The Tc99 had already reached the caecum in large quantities before the H2 PPM level has reached the threshold for an abnormal test. This demonstrates that the increased H2 production results from fermentation by colonic bacteria, not by abnormal bacteria small intestine (ie, SIBO).94
Differences in the microbiota in FBD and the link to pathophysiology
There is little known about the small intestinal microbiota as the small intestine is relatively inaccessible (summarised in table 2; box 2).75–85 Culture studies show considerably fewer bacteria compared with the colon with a marked gradient from duodenum to distal ileum. The bacteria are typically Gram-positive aerobes proximally and Gram-negative and Gram-positive anaerobes and facultative anaerobes in the terminal ileum. Culture-independent studies of the small intestinal microbiota are in their infancy but suggest complexity not appreciated by standard culture techniques, including marked individual differences, fluctuations over time (even within the same day), age-related differences and several phylotypes not previously identified.86–88 Moreover, a recent paper indicated that the small intestinal microbiota are driven by a rapid uptake and conversion of available simple carbohydrates in which Streptococcus spp. play an important role.89
Summary of studies culturing small bowel microbiome
Box 2 Relevance of studies showing changes in microbiota in irritable bowel syndrome
-
The relevance of small intestinal bacterial overgrowth in irritable bowel syndrome (IBS) remains unclear due to methodological problems, influence of confounding factors and large differences between studies.
-
Heterogeneity of IBS and variation in methods used to study the faecal microbiota have resulted in conflicting reports of differences from healthy controls.
-
The microbiome may contribute to IBS symptoms by altering gut neuromotor-sensory function, barrier function and/or the brain–gut axis.
The role of SIBO in the pathogenesis of IBS is very controversial because the breath tests employed to establish this role have not been validated.90 ,91 Even the validity of the ‘gold standard’, jejunal cultures >105 cfu/ml with colonic-type bacteria, has been challenged, largely because this cut-off was established from samples following surgical diversion.91 Studies in IBS patients showed relatively few bacteria in the duodenum and proximal jejunum and no obvious differences from controls (table 2). Preliminary studies suggest that more IBS patients have SIBO when a lower cut-off of >103 cfu/ml is used but well-designed studies are needed.82 ,85 Available molecular studies are not adequately designed to establish whether SIBO is involved in IBS but have significant potential.
Several confounding factors, including acid suppression by proton pump inhibitors (PPIs) and altered motility, have been implicated in the studies of SIBO and IBS.92–94 Some studies suggest that PPI use might lead to symptomatic SIBO or at least increased numbers of bacteria and that following antibiotics they accelerate recurrence, but this depends on the tests employed and criteria applied.95 Although the link between SIBO and IBS is largely based on breath testing, most positive lactulose breath tests reflect rapid transit to the caecum rather than true SIBO94 (figure 4). Other factors such as antibiotics, probiotics and prebiotics, and other dietary items such as FODMAPs could also influence microbiota in IBS patients and result in a potentially spurious association.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Plot chart of currently available strategies for modifying gut microbiota aiming to demonstrate the relationship between the effectiveness and invasiveness/safety of the proposed approach. FODMAP, fermentable oligo-, di- and mono-saccharides and polyols; PPI, proton pump inhibitor.
Earlier culture-based assessment of faecal microbiota obtained from patients with IBS demonstrated decreased faecal lactobacilli and bifidobacteria, and increased facultative bacteria dominated by streptococci and Escherichia coli as well as higher counts of anaerobic organisms such as Clostridium. 96 ,97 Studies using molecular-based techniques reveal changes in faecal microbiota composition in IBS versus controls (table 3). Interestingly, a recent study demonstrated that faecal microbiota of IBS patients could be grouped in a cluster which was completely different from that of healthy controls.114 Nonetheless, results to date are inconsistent and sometimes contradictory (table 3). This may reflect differences in molecular techniques employed, the use of single samples that are not linked to fluctuating symptoms (especially as studies suggest IBS faecal microbiomes are less stable), and probably other factors such as diet and phenotypic characterisation of patients. In addition, it should be realised that faecal samples do not necessarily reflect other parts of the GI tract.
Summary of culture and molecular studies of colonic microbiome
The finding that IBS can develop following infective gastroenteritis prompted studies evaluating the role of inflammation in IBS, but there are fewer studies that focus on the associated changes in gut microbiota, which might be just as significant. Infective gastroenteritis produces a profound depletion of the commensal microbiota,118 whose production of metabolites such as SCFAs and antibiotics normally inhibits pathogen colonisation, as can be seen from the loss of colonisation resistance after antibiotics.119 It is unclear just how completely and over what time span recovery occurs.
Infective gastroenteritis is common, with an incidence of 19/100 person years in the UK.120 A third of episodes are viral (Norovirus/Rotavirus being the commonest). The commonest bacterial infections, Campylobacter and Salmonella, account for 10% and 3%, respectively. Onset of new IBS symptoms after a bout of infective gastroenteritis is relatively common, reported by 6%–17% of IBS patients,121 while a recent internet survey reported 18%,122 with around 40% beginning while travelling. The clinical features of post-infectious-IBS are predominantly those of IBS-diarrhoea (IBS-D).123 ,124 A recent meta-analysis pooling 18 studies indicated a relative increased risk of developing IBS 1 year after bacterial gastroenteritis (mostly Shigella, Campylobacter and Salmonella), RR=6.5 CI (2.6–15.4), an effect still apparent at 36 months, RR=3.9 (3.0–5.0).125 Viral gastroenteritis, in keeping with the lesser tissue injury, shows a reduced incidence of post-infectious-IBScompared with bacterial infections126 ,127 in which the strongest risk factors are bacterial toxicity,128 prolonged duration of diarrhoea,124 rectal bleeding129 and fever.125 Acute enteritis is associated with a prolonged increase in mucosal cytotoxic T lymphocytes and increase in enteroendocrine cells.123 Other studies have shown the importance of increased 5HT containing cells in IBS-D130 and increased sensitivity in IBS-D with increased EC cell counts,131 accelerated gut transit and visceral hypersensitivity.132 These effects on gut physiology will impact on the gut microbiota environment. An early study of children with acute gastroenteritis demonstrated alkalinisation of stool pH, likely due to the decrease in bacterial metabolites (SCFAs) and a fall in numbers of Bacteroides, Bifidobacterium, Lactobacillus and Eubacterium.133 Conventional enumeration of faecal bacteria showed a 10-fold fall in anaerobes (Bacteroidaceae and Eubacterium), little change in aerobes, but 109 cfu/g of pathogens. Another study using conventional culture methods showed a reversal of the normal anaerobe/aerobe dominance during acute infection.134 More recent human studies using modern culture-independent methods tended to confirm these findings.135 ,136 PCR-denaturing gradient gel electrophoresis profiling of 16S rRNA genes showed a reduced diversity, often associated with a dominant band suggesting overgrowth of one subtype, which may not always be the original pathogen. A recent clinical trial of an oral rehydration solution containing a prebiotic, amylase resistant starch in acute diarrhoea in India, including children aged 3 months to 5 years, used PCR primers directed at selected bacteria, for example, Eubacterium spp. and Faecalibacterium prausnitzii, key bacteria involved in starch fermentation. These studies showed a decline in some anaerobes (Bacteroides spp., Eubacterium spp. and Faecalibacterium prausnitzii) while other genera including Bifidobacterium spp. were unchanged.135 This depletion of anaerobes could be due to acceleration of transit, which could lead to a loss of the anaerobic niche. Since these are the key bacteria involved in colonic salvage of unabsorbed carbohydrate,137 this may also contribute to the diarrhoea phenotype by preventing fermentation to SCFAs, which are known to stimulate colonic salt and water absorption, both directly and by inducing increased expression of transporters.138–140 Previous earlier studies in IBS-D suggest impaired SCFA concentrations and production rates in ex vivo incubation, which may also reflect reduced anaerobes.141
Another cause of depletion of anaerobes is broad-spectrum antibiotics. There are no RCTs, but epidemiological studies show an association between antibiotic use and an increased risk of PI-IBS.142 A study of children showed that 3 months after Salmonella infection, vomiting, abdominal pain and diarrhoea were reported by 9.5% of those treated with antibiotics but only 2.9% of those who received no antibiotics.142
Changes in the interaction between intestinal microbiota and host factors (eg, age, diet, transit, host genetic factors, antibiotics) could be important for IBS pathophysiology. These factors, in turn, could be related to changes in homeostatic pathways including barrier function, neuromotor sensory function and the brain–gut axis.143 ,144 For example, bidirectional signalling between the microbiota and the epithelium regulates epithelial secretion of mucus as well as other defence factors involved in regulating the microbiota. Changes in these factors (eg, changes in mucus layer and increased β-defensin-2 peptide) have been detected in patients with IBS and functional diarrhoea and suggest a microbiota–host immune system engagement.145 ,146 In line with this concept, there is also recent demonstration that IBS patients have increased colonic mucosal expression of receptors recognising specific microbiota-related substances (such as Toll-like receptor-4 which recognises bacterial lipopolysaccharides)147 or increased titres of circulating antibodies against components of the indigenous microbiota (ie, antiflagellin antibodies).148 Several studies demonstrated low-grade activation of innate and adaptive mucosal immune response in large subgroups of patients with IBS.12 ,149 Increased activated mast cells, CD3+ve, CD4+ve and CD8+ve T cells have been detected in both postinfectious IBS and non-specific IBS.12 ,149 The relative importance of mast cells in this setting is demonstrated by the abundance of this immune cell type over other immunocytes and by increased release from mucosal biopsies of histamine, tryptase and prostaglandins.150 ,151 Mast cells were located in closer vicinity to mucosal innervation and correlated with the severity and frequency of abdominal pain in patients with IBS.152 There are potential implications of mucosal immune activation for sensorimotor dysfunction of patients with IBS. Histamine and tryptase released from mucosal biopsies of patients with IBS evoked increased mesenteric sensory afferent activation and induced visceral hyspersensitivity via histamine-1 receptors and proteinase activated-2 receptors when applied to recipient rats.150 ,151 Intestinal microbiota may well be an active participant in this scenario through stimulation of the immune system,153 likely in the subgroup of subjects showing increased epithelial permeability which could154 expose the immune system to an abnormal microbial antigenic load. Overall, the results suggest that bacterial–host interactions may be initiated by components of the microbiota that can cross the mucus and adhere to epithelial cells, inducing activation of the mucosal innate defence system even in the absence of mucosal destruction.
The use of probiotics, particularly in animal models, also demonstrates that their secreted products or metabolites can modulate contractility of intestinal smooth muscle and visceral sensitivity.59–61 Moreover, application of probiotics can recover neuromotor-sensory dysfunction in IBS-like models.
Modulation of the brain–gut axis is particularly relevant in IBS because psychological comorbidity is common. Some forms of psychological stress in animal studies can induce shifts in the bacterial composition of the gut that is accompanied by systemic cytokine response and increased intestinal permeability.155 The interplay may be bidirectional as suggested by animal studies showing that the microbiota can affect brain chemistry and behaviour.156 Nonetheless, for the time, the potential relevance of brain–microbiota interactions have yet to be shown in humans in general and in FBD in particular.
GI disorders mimicking and overlapping with FBDs
Although celiac disease, IBD or diverticulitis can coexist with IBS, an ‘IBS’ diagnosis in the presence of an organic disease may be challenging.
Gluten causes coeliac disease in genetically susceptible people and causes gut dysfunction in mice and can generate IBS symptoms in the absence of coeliac disease.157 Some patients with IBS lack tissue transglutaminase antibodies or histological markers of coeliac disease yet still respond symptomatically to a gluten-free diet. This entity is termed ‘non-coeliac gluten sensitivity’ or ‘gluten sensitive IBS’.157–159 The underlying mechanisms in humans remain unclear. Mouse models indicate that gluten can induce activation of innate immunity, increased small intestinal permeability,160 neuro-muscular dysfunction159 and dysbiosis161 in the absence of autoimmunity.
IBS-like symptoms are common in IBD patients in long-standing remission, or are frequently reported in patients before the diagnosis of IBD.162 ,163 It is possible that IBS and IBD coexist with a higher than expected frequency, or may exist on a continuum, with IBS and IBD at different ends of the inflammatory spectrum. A study investigating IBS symptoms in IBD patients who were thought to be in clinical remission demonstrated high levels of calprotectin levels; this suggests that in most cases IBS symptoms are the result of undetected ongoing inflammation.164 Underlying mechanistic links are lacking but it is tempting to raise the hypothesis that the intestinal microbiota may be a common factor in both diseases.165 In fact, as with IBS (tables 2 and 3), faecal166–171 and mucosal-associated dysbiosis167 ,172–178 has been described IBD.
A high proportion of patients hospitalised with acute diverticulitis continue to have persistent symptoms that mimic IBS179 despite the absence of complications.180 Some uncontrolled studies claim benefit from antibiotics and/or mesalazine suggesting a role for the microbiota in this syndrome.181
Treatment implications: antibiotics, probiotics, prebiotics and synbiotics
As the microbiota may be disturbed in functional GI disorders, a potential treatment approach is to try to correct dysbiosis either by the administration of an antibiotic or a preparation of ‘beneficial’ bacteria (box 3).
Box 4 Diagnostic and therapeutic general recommendations
-
There is currently no clinically useful way of identifying whether the microbiota are disturbed in particular patients with irritable bowel syndrome (IBS).
-
Dietary evaluation and exclusion of possible sources of unabsorbable carbohydrates including fermentable oligo-, di- and mono-saccharides and polyols and excessive fibre could be beneficial in select patients.
-
Probiotics have a reasonable evidence base and should be tried, for a period of at least 1 month, at adequate doses before a judgement is made about the response to treatment.
-
The utility of testing for small intestinal bacterial overgrowth (SIBO) in the setting of IBS remains an area of uncertainty.
-
If SIBO is strongly suspected based on clinical presentation and testing is being considered, using stringent criteria for the glucose breath test or jejunal aspirate appear to be the best tests.
-
Consideration should be given to discontinuing proton pump inhibitors in those with SIBO.
-
There is emerging evidence that non-absorbable antibiotics may have the potential to reduce symptoms in some patients with IBS.
Antibiotics
Despite evidence that previous antibiotic use may be related to the development of IBS,182 ,183 and the fact that antibiotic treatment may increase the development of long-term digestive symptoms after bacterial gastroenteritis,142 poorly absorbable antibiotics might still have therapeutic potential in this condition.184 Neomycin was the original choice184 ,185 although interest is now focused on a non-absorbed derivative of rifampicin called rifaximin.186
There are three fully-published, double blind, placebo controlled trials of rifaximin in FBD187–189 and the data suggest an improvement in symptoms, especially bloating and flatulence for approximately 10 weeks following treatment187 ,189 with a therapeutic advantage over placebo around 10%. The doses used in these and other studies vary between 600 and 2400 mg daily for 7–14 days190–195 but there remain concerns about antibiotic resistance and possible Clostridium difficile infection although so far these issues have not appeared to be a problem.196–200
Thus, a short course of gut-specific antibiotics may have utility in some patients with IBS but we need to know more about predictors of treatment responsiveness, antibiotic resistance, the efficacy and safety of re-treatment schedules as well as the optimal dosing regimen.201 ,202
Probiotics
Probiotics are live microorganisms which when administered in adequate amounts confer a health benefit on the host203 with the most commonly used being the lactobacilli and bifidobacteria. Probiotics can be packaged in many formulations containing just one organism or a mixture and have a wide range of activities with evidence supporting an effect on at least some of the putative pathophysiological mechanisms implicated in IBS, such as visceral hypersensitivity,59 ,60 ,204 ,205 GI dysmotility,206–210 intestinal permeability,204 ,211 ,212 the intestinal microbiota213 ,214 and immune function215 although these effects can differ considerably between one organism and another. Thus, just because one organism is beneficial, this does not mean that related organisms will behave similarly. For use in gastroenterology, it is important that a preparation contains sufficient quantities of microbes which need to be acid and enzyme resistant with good mucosal adherence also being an advantage.
Table 4 lists the results of the fully published placebo controlled probiotic trials to date.208–210 ,212 ,214–240 Unfortunately, their designs vary considerably;241–244 some of the older studies are of poor quality, and few attempt to define the mechanism of action or assess whether symptomatic improvement is accompanied by a change in the microbiota. A recent systematic review reported that studies of poorer quality tended to show larger effects and published data indicate a publication bias, with non-reporting of negative effects in small trials.244 Around three-quarters of these studies were positive, of which four were in children, although different symptoms improved and the therapeutic gain over placebo was generally modest. Furthermore, it remains unclear which organisms are most effective as, for instance, some mainly reduce bloating and flatulence,209 ,210 ,218 whereas others improve bowel frequency,222 and some have a positive effect on global symptom scores.214 ,215 ,224 ,226 ,229 ,230 ,233 In some of the better quality trials bifidobacteria, such as Bifidobacterium infantis 35624,215 ,224 ,241 Bifidobacterium lactis DN 173010208 ,225 and Bifidobacterium bifidum MIMBb75,226 seem to be advantageous and in others probiotic mixtures appear to be useful.214 ,229 ,233 In only one study was there symptom deterioration221 although some large, high quality trials have been negative.221 ,231 ,234 ,235
Placebo controlled clinical trials of single or mixed probiotic preparations in IBS
Diet, fibre, prebiotics and synbiotics
There are few proper randomised, placebo controlled trials of diet modification because of the difficulty in controlling for the placebo effect. One randomised controlled trial showed bran aggravated symptoms;245 excluding bran should help, and many patients believe this is true.246 A prebiotic is a product that, on ingestion, stimulates the growth of beneficial bacteria already present in the host, which promotes the health of the individual.247 ,248 A variety of oligosaccharides serve this function and a synbiotic is a combined prebiotic and probiotic. One of the earliest prebiotics was lactulose, an unabsorbable disaccharide laxative that increases the faecal concentrations of Bifidobacterium spp.45 ,249 as does inulin which, like lactulose, increases flatulence47 and thus makes it unlikely it will help IBS patients.
To date, there has only been one double blind, placebo controlled trial of a prebiotic in IBS which used a trans-galactooligosaccharide mixture.250 Compared with placebo this prebiotic reduced symptoms and stimulated the growth of bifidobacteria but clearly more research is required on dosing and the relative merits of other compounds. With regard to synbiotics, there are some studies but their design is not sufficiently robust to draw any firm conclusions251–255 although the concept of combining a prebiotic and probiotic is theoretically attractive. Thus, attempting to modify the microbiota in patients with functional GI disorders shows some promise. However, we need to know how symptomatic improvement is achieved: is it mirrored by a change in gut microbiota or is some other mechanism involved?
Clinical guidance regarding modulation of intestinal microbiota in IBS
While the science regarding the role of microbiota in FGIDs remains in its infancy, patients are exposed to conflicting claims concerning the symptomatic benefit from modulating gut microbiota. This section aims to help clinicians give the best advice, despite limited evidence (box 4).
Diet profoundly alters the microbiota. Reducing intake of fibre256 or FODMAPs257 is one of the simplest and safest ways of altering gut microbiota, which can lead to improvement in bloating and diarrhoea, an effect which may last for years.258 However, so far the evidence to support widespread use of FODMAP reduction in patients with IBS is limited and comes mainly from one research group. Systematic exclusion diets may also help258 but are laborious; targeted exclusion of regularly consumed suspects, such as dairy, wheat, fruit and vegetables, may be more practical.
The safety of probiotics in IBS is acceptable but some aggravate symptoms221 and so patients should be warned of this possibility. At present, the strongest evidence is for Bifidobacterium infantis 35624 at a dose of 1×108 cfu/day taken for at least 4 weeks.224 It remains unclear who benefits from which variety of probiotic since there are many incompletely answered questions surrounding this therapeutic approach, including:
-
Are single organisms better than mixtures or vice versa?
-
Do some mixtures of organisms contain strains that are competitive or antagonistic without additive effects?
-
Can probiotic foods and drinks be administered simultaneously?
-
What are the best delivery systems—liquids or capsules?
-
How can viability and bioavailability be ensured?
-
What are the optimal dosing regimens and their duration?
-
What is the frequency of host colonisation?
-
Probiotics are not potent pharmacological agents: what patient group should be targeted?
-
Are there any groups of patients where probiotics might be contraindicated such as newborns, immunocompromised or seriously ill individuals?
-
Are there any safety issues about some strains of probiotics, for example, those of Escherichia coli?
-
Should different probiotics be given to specific subgroups of IBS patients?
-
Which symptoms of IBS should be the main target for therapy?
-
What are the possible mechanisms behind symptom improvement?
-
How should doctors and patients be advised about their administration?
The considerable acid suppression induced by PPIs may alter upper gut microbiota and can potentially induce IBS symptoms.92 Thus, it is worth considering PPI discontinuation in selected IBS patients on PPIs for unclear reasons, especially if their symptoms started with PPI therapy.
The most direct way of altering gut microbiota is to use broad-spectrum antibiotics. However, rapid development of antibiotic resistance leads to concerns about using antibiotics in such a ubiquitous and chronic condition. Moreover, it is likely that patients may require repeated courses of therapy, as trial evidence suggests the benefit diminishes by 12 weeks.189 The best evidence is for rifaximin 550 mg, thrice daily for 2 weeks.189 The number-needed-to-treat was 11 which should be compared with 4 for ‘placebo without deception,259 7 for alosetron,260 8 for linaclotide261 and 14 for tegaserod.262
Although rifaximin appears to be well tolerated and safe, given its relatively low potency its use should be restricted to difficult cases since its widespread use could promote resistance, such as rifampin-resistant strains of stapyholococci.196 Figure 4 provides a plot chart of currently available strategies for modifying gut microbiota according to the effectiveness and invasiveness of the proposed approach, and general recommendations appear in box 4.
Conclusions and recommendations for future research and development
Although there is good evidence supporting the concept that the intestinal microbiota is perturbed in patients with FBD, we still lack data on the mechanisms through which host–microbiota interactions underlie pathophysiology and generate symptoms; we need to overcome several boundaries that hold back our knowledge in this field.
The SIBO hypothesis in IBS remains a matter of debate because the breath tests and the small bowel culture techniques have not been validated. In addition, confounding factors, including the use of antibiotics or PPIs, have not been taken into account in many studies. The wide heterogeneity of FBD and the inter-individual variability of microbiota profiles suggest that larger sample size studies (both in the pathophysiology and therapeutic settings) are of key importance in the future. Attention should be directed to the assessment of correlations between microbiota changes with patient's symptoms. Whenever possible, studies should be stratified by factors known or assumed to affect intestinal microbiota (eg, age, diet, enterotype) and designed to reduce potential confounding factors (eg, antibiotics, probiotics, laxatives, prokinetcs, PPI and mesalazine). Although faecal samples are relatively easy to obtain, future work should better characterise microbial populations at the luminal and mucosal level which may differ substantially from faecal microbiota. Host–microbiota interactions are dynamic events and likely influenced by several factors. This suggests that there is a need for longitudinal studies assessing gut microbiota during remission, and symptom flare-ups, stress, infection or following dietary manipulation and the use of probiotics, prebiotics and antibiotics. The effect of bowel transit on microbiota profiles and correlation with symptoms should also be assessed.
One important limitation of available studies is their descriptive rather than mechanistic nature. Accordingly, studies should be directed at clarifying cause–effect relationships between microbiota changes and bowel dysfunction. In this way, microbiota signatures can be developed to help identify IBS biomarkers which might, in turn, offer therapeutic targets. For example, the theory that luminal bacteria may drive low-grade intestinal immune activation should now be substantiated by mechanistic and interventional studies. Data in rodents suggest the existence of a bidirectional interplay between the brain and gut microbiota.144 In addition, the existence of systemic immune responses to microbial luminal antigens (antiflagellin antibodies)148 provides initial evidence that microbial homeostasis may be perturbed beyond the GI tract. These aspects need to be further explored to open new avenues of research in FBD.
Currently, there are promising results suggesting that a subgroup of patients with FBD may respond favourably to a short course of gut-specific antibiotics. However, most probiotic and antibiotic trials are underpowered and suffer suboptimal design. Bloating and flatulence appear to be especially responsive to non-absorbable antibiotics. In order to safely direct these treatment options to the appropriate patients, we need to know more about predictors of treatment responsiveness, the risk of development of antibiotic resistance, the efficacy and safety of re-treatment schedules, and the optimal dosing regimen.201 ,202 Further studies should also investigate the mechanism and site of action of non-absorbable antibiotics since amelioration of gas-related symptoms in patients occurred also in patients with no evidence of SIBO.188 Probiotics seem to have a positive, albeit modest, effect in both children and adults with FBD, especially IBS. However, head-to-head comparisons between different probiotic products would be useful and future trials need to be large scale, high-quality and use valid end points. Trials should also explore the mechanisms behind symptom improvement.
Faecal transplantation is efficacious in 145⁄166 (87%) patients with fulminant and refractory C difficile infection. This procedure has also been proposed for the treatment of IBS but further research is needed.263
In conclusion, a better definition of the role of intestinal microbiota in the pathogenesis and pathophysiology of FBD represents a challenge for the future. Although promising, therapeutic implications will need to be better defined in well-conducted, large clinical trials. A strict cooperation of experienced clinical researchers with microbial ecologists should be considered an important factor for the success of these future studies.
Acknowledgments
The authors would like to thank Professor Willem de Vos, Professor Eamonn Quigley, Professor Patrizia Brigidi and the Rome Foundation board members for critical revision of the manuscript. We also acknowledge the valuable help of Cesare Cremon in manuscript editing and the expert secretarial support from Emma Bradley.
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Box 3 Modulation of intestinal microbiota in functional bowel disorders
-
A short course of a non-absorbable antibiotic such as rifaximin has been shown to moderately improve the symptoms of irritable bowel syndrome (IBS), particularly bloating and flatulence. Improvement persists after the cessation of treatment but the exact duration of this effect remains uncertain.
-
The majority of trials of probiotics in IBS show some degree of efficacy although some of the early studies were of very poor quality.
-
Prebiotics and synbiotics should theoretically have the potential in treating functional gastrointestinal disorders but there are as yet no reliable data to support this view.
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data Supplement 1 - Online Supplement 1
Footnotes
-
Funding This work is supported by Rome Foundation, USA. In addition, the authors would like to acknowledge the following funding sources: Italian Ministry of Education, University and Research (PRIN 2009); Fondazione Cassa di Risparmio and IMA, Bologna, Italy (GB); Scottish government RESAS support (HJF); The Swedish Medical Research Council (grants 13409, 21691 and 21692), the Marianne and Marcus Wallenberg Foundation, and the University of Gothenburg, Centre for Person-Centred Care (GPCC), Sahlgrenska Academy, University of Gothenburg and by the Faculty of Medicine, University of Gothenburg (MS); Shire/Movetis, Amgen, Ironwood (BS); Canadian Institute of Health Research (CIHR) and Crohn's and Colitis Foundation of Canada (CCFC) (SV); Lesaffre International, Norgine, National Institute for Health Research Biomedical Research Unit Grant (RS); CAG/CIHR, CCA (Canadian Celiac Association), CCFC and Nestec (EFV); EFV holds a Career Award from the Department of Medicine, McMaster University.
-
Competing interests This is a Rome Working Team Report. All authors are responsible for writing the study interpretation of data, and critical revision of the manuscript. The authors would like to disclose the following potential competing interests: Alfa-Wasserman, Prometheus, Shire/Movetis, Sofar (GB); Danone Research, Arla Foods, Novartis, Shire/Movetis, AstraZeneca (MS); Ironwood, Shire/Movetis, Prometheus (BS); Boeringer Ingelheim & Ironwood (RS); Ferring Canada and US (SV); Nestec grant support (EFV); Novartis Pharmaceuticals, GlaxoSmithKline, Solvay Pharmaceuticals, Pfizer Global Research and Development, Rotta Research, Proctor and Gamble, Danone Research, Astellas Pharma, Ironwood Pharmaceuticals, Sucampo Pharmaceuticals, Almirall Pharma, Movetis UK, Norgine, Chr Hansen, Boehringer-Ingelheim, and Heel GMBH (PW).
-
Provenance and peer review Not commissioned; externally peer reviewed.