Article Text

Abstract
Objectives To compare the appendiceal microbiomes and examine the prevalence of Campylobacter species in the appendices of adult subjects with confirmed acute non-perforated appendicitis and controls with healthy appendices.
Design Archived samples of formalin-fixed paraffin-embedded appendiceal tissues were obtained from 50 consecutive female subjects who underwent appendectomy for acute, non-perforated appendicitis, and 35 consecutive female controls who underwent incidental appendectomy during gynaecological surgery.
Results 16S rRNA gene sequencing revealed that the relative abundances (RAs) of the major phyla in appendiceal tissues (Firmicutes, Proteobacteria, Bacteroidetes, and Actinobacteria) were similar in both groups. Beta diversity was significantly different due to differences in Bacteroidetes and Proteobacteria (p<0.0001). Within Proteobacteria, RAs of classes Alphaproteobacteria (~21%, fold change (FC)=1.31, false discovery rate (FDR) p value=0.03) and Epsilonproteobacteria (~1%, FC=0.25, FDR p value>0.05) were increased in acute appendicitis samples. RAs of unknown genera from families Burkholderiaceae and Enterobacteriaceae were decreased in appendicitis samples, and 14 genera were increased, including Neisseria, Acinetobacter and Campylobacter. Quantitative PCR revealed that levels of Campylobacter jejuni DNA, but not other Campylobacter species or pathogens tested, were significantly higher in appendicitis samples than in controls (p=0.013). Using a cut-off of 0.31 pg/µL, 40% of appendicitis cases and 6% of controls were positive for C. jejuni, indicating specificity of 93.7% (95% Cl 79.2 to 99.2), sensitivity of 40.9% (95% Cl 24.7 to 54.5), and OR of 10.38 (Fisher’s p value=0.0006, 95% Cl 2.3 to 47.4).
Conclusions Our findings indicate that Campylobacter jejuni may be a significant cause of acute appendicitis. This supports earlier studies and suggests that targeted antibiotic therapies could be an alternative treatment for a subset of non-complicated acute appendicitis cases.
- appendicitis
- campylobacter jejuni
- enteric bacterial microflora
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Summary box
What is already known about this subject?
Acute appendicitis is the most common cause of abdominal surgery.
There is little data exploring the microbiome of the appendix in healthy individuals and in appendicitis.
The few studies of the appendiceal microbiome are mostly in paediatric populations.
What are the new findings?
Although the relative abundances (RAs) of major phyla are similar in inflamed appendiceal tissues from adult subjects with acute appendicitis versus non-inflamed tissues from healthy appendices, there is a significant difference in beta diversity between the two groups, driven primarily by differences in Bacteroidetes and Proteobacteria.
Within Proteobacteria, RAs of many taxa were significantly altered, including increases in classes Alphaproteobacteria and Epsilonproteobacteria, in 23 families including Moraxellaceae, Pasteurellaceae, Neisseriaceae and Campylobacteraceae, and in 14 genera including Neisseria, Acinetobacter and Campylobacter.
Levels of C. jejuni, but not other Campylobacter species or gastrointestinal pathogens tested, were significantly higher in appendicitis samples than in controls.
How might it impact on clinical practice in the foreseeable future?
Our findings confirm previous reports suggesting that C. jejuni may be a significant cause of acute appendicitis.
This also supports recent data that targeted antibiotic therapies may be an alternative treatment for a subset of non-complicated acute appendicitis cases.
If C. jejuni in the appendix predicts an antibiotic responsive condition, this would better stratify subjects with acute appendicitis.
Introduction
Acute appendicitis is defined as an inflammation of the appendix, and is associated with symptoms including right lower abdominal pain, nausea, vomiting and fever. The most significant complications are sepsis and perforation,1 2 and appendectomy remains the most common abdominal surgical emergency in the world.3 Over 280 000 appendectomies are performed annually in the USA alone,2 with associated US healthcare costs estimated at over 3 billion dollars per year.3 The majority of cases have classically been thought to result from obstruction of the lumen of the appendix (such as may be caused by stool, a fecalith, or lymphoid hyperplasia), leading to inflammation and bacterial overgrowth. However, more recent research has focused on the potential roles of overgrowth of appendiceal microbes and/or infection with bacterial pathogens.4–9
Since McBurney performed the first appendectomy in 1889,10 surgery has remained the gold standard for treatment of acute appendicitis. However, the clinical diagnosis can be challenging, and many of the conditions within the differential diagnosis require other treatment strategies. Even if the diagnosis of appendicitis is confirmed, there remains a question as to whether surgery is needed acutely. The negative appendectomy rate remains high, and there is some debate as to whether the use of computed tomography (CT) impacts this.11 12
Given the recently identified roles for bacteria as potential causes of appendicitis, it is not surprising that a number of reports suggest there may be a role for antibiotic treatment in acute uncomplicated appendicitis.13–15 In fact, a recent randomised clinical trial of 257 patients treated with antibiotics for uncomplicated acute appendicitis found the cumulative incidence of recurrent appendicitis to be 27.3% at 1 year, 35.2% at 3 years and 39.1% at 5 years, and concluded that antibiotic treatment may be an alternative to surgery in as many as 60% of cases.16 However, it has also been suggested that the appendix may function as a reservoir for commensal bacteria, allowing for reseeding the gut microbiome following perturbations such as antibiotic treatment, severe diarrhoea, and so on.17 This function could be impacted by the use of broad-spectrum antibiotics, but would clearly also be impacted by appendectomy.
A challenge in defining bacteria which may contribute to appendicitis is that, as for the gut microbiome, the ‘healthy’ appendiceal microbiome remains poorly understood. Earlier studies attempted to identify causative bacteria using culture-based techniques,18 19 fluorescence in situ hybridisation,4and so on, several of which suggested potential links between Campylobacter species and appendicitis.20–22 More recent studies examined the appendiceal microbiome using next-generation sequencing,5–9 the majority using paediatric populations.6–8 Consistent findings included increased abundance of phylum Fusobacteria,7 genus Fusobacterium6 and specific Fusobacterium spp8 in subjects with appendicitis, as well as increases in genera typically associated with the oral cavity, such as Porphyromonas and Parvimonas.6–8 In this study, we used 16S rRNA gene sequencing and quantitative PCR (qPCR) to compare the appendiceal microbiomes and examine the prevalence of Campylobacter species in the appendices of adult subjects with confirmed acute non-perforated appendicitis and control subjects with healthy appendices.
Methods
Samples and study subjects
Archived samples of formalin-fixed paraffin-embedded (FFPE) appendiceal tissues were obtained from (1) 50 consecutive female subjects who presented to a tertiary care institute with acute non-perforated appendicitis and underwent appendectomy during the same admission; and (2) 50 consecutive control subjects without acute appendicitis who underwent incidental appendectomy during gynaecological surgery for indications such as endometriosis and leiomyoma during the same time period. Samples of FFPE appendiceal tissues were reviewed by an experienced pathologist to confirm the diagnosis of acute appendicitis and identify areas of inflammation in the appendicitis group, and to confirm the absence of inflammation in the control group. Subjects with perforated appendicitis, known bowel disease (coeliac disease, inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS)), concomitant active Clostridium difficile colitis or who had taken antibiotics within the 3 months prior to surgery were not included in this study.
Patient and public involvement
Patients and the public were not involved in the design of this study.
Sample processing
Sample identification
Tissue with all layers of appendix wall containing areas of inflamed tissue in sections of FFPE appendiceal tissues from subjects with acute appendicitis were identified by a pathologist. Inflamed tissue was defined as acute inflammation of any layer of the appendix wall (the mucosa, submucosa, muscularis mucosa, serosa, mesoappendix). To do this, the first four 8 µm sections of the FFPE tissue block were discarded in order to reach tissues unexposed to oxygen. A set of nine further tissue sections were then cut from the block and mounted on slides. The first slide from each set was stained using H&E and the area of inflammation was circled. The eight sequential unstained slides from each subject were reserved for DNA isolation (detailed below). Similar histological analysis was performed on FFPE healthy appendiceal tissues from control subjects, and tissues with all layers of appendix wall free of any sign of inflammation were marked on the H&E-stained slides. The eight sequential unstained slides from each subject were reserved for DNA isolation.
DNA isolation
Microbial DNA isolation was performed using tissues from unstained slides only, using QIAamp DNA FFPE Tissue Kits (Qiagen, Hilden, Germany) and following a protocol that was optimised in our laboratory. Specifically, using the H&E-stained slide as a guide, the area of interest only was carefully removed from each of four sequential unstained slides by microdissection with a sterile razor, in order to prevent cross-contamination with other tissue areas from the same slide. These microdissected tissue samples were then transferred to a sterile 2 mL tube. One millilitre of xylene was immediately added to the tube and vortexed vigorously for 10 s. The samples were centrifuged at full speed (20 000×g) for 2 min at room temperature and the supernatant was removed by pipetting, taking care not to disturb the pellet. One millilitre of ethanol (96%–100%) was added to the pellet and mixed by vortexing for 3–5 s. After centrifugation at full speed for 2 min at room temperature, the supernatant was carefully removed, again taking care not to remove any of the pellet. The ethanol residue was then allowed to evaporate at room temperature for 10 min.
The pellet was resuspended in 360 µL of Buffer ATL (Qiagen) and 40 µL proteinase K and mixed by vortexing. After incubating at 56°C for 30 min and then at 90°C for 10 min, tubes were brought to room temperature and 4 µL of RNase A (100 mg/mL) (Qiagen) and garnet beads (0.70 mm) (Qiagen) were added to each tube. Samples were then vortexed horizontally for 10 min. Tubes were centrifuged at room temperature for 6 min at 4500×g, and the supernatant (approximately 200 µL) was transferred to a 2 mL sterile tube. Then, 200 µL of Buffer AL (Qiagen) was added to the sample and mixed thoroughly by vortexing. Next, 200 µL of ethanol (96%–100%) was added and mixed again by vortexing. The tubes were briefly centrifuged to remove drops from the inside of the lid and the entire volume was transferred to a QIAamp MinElute column. The QIAamp MinElute columns were centrifuged at 6000×g (8000 rpm) for 1 min, and after placing the column in a clean 2 mL collection tube, two wash steps were performed with Buffer AW1 and Buffer AW2 (Qiagen) according to the manufacturer’s protocol. After the wash steps, the membranes were dried completely by centrifuging the columns within new 2 mL collection tubes for 3 min at full speed. The DNAs were then eluted with 60 µL of Buffer ATE (Qiagen). DNAs were quantified using Qubit ds DNA BR Assay kits (Invitrogen by Thermo Fisher Scientific, Waltham, Massachusetts, USA) on a Qubit 4 Fluorometer (Invitrogen).
Library preparation and 16S rRNA sequencing
16S library preparation for DNAs from all groups was performed per our published protocol.23 An optimised clean-up step was performed with Agencourt AMPure XP beads using modifications proposed by Quail et al,24 and 5 µL of the final Amplicon PCR product was used for the Index PCR, which was performed using the Nextera XT Index kit and 2 x KAPA HiFi HotStart Ready Mix, following the Illumina protocol. After a second modified clean-up step, the final product was quantified using Qubit ds DNA BR Assay kits and Qubit 1X dsDNA HS Assay kits on a Qubit 4 Fluorometer, and analysed using Agilent DNA 1000 chips (Agilent Technologies, Santa Clara, California, USA) and Agilent HS DNA chips (Agilent) on an Agilent 2100 Bioanalyzer System.
16S rRNA sequencing
The V3 and V4 libraries prepared using DNA from appendicitis samples and control samples were sequenced using a MiSeq Reagent Kit v3 (600-cycles) on a MiSeq System (Illumina, San Diego, California, USA) as described previously.23 Further, 2×301 cycles of paired-end sequencing were performed according to manufacturer’s protocol and 5%–10% Phix (Illumina) was added to each library pool.
Sequencing analysis
Operational taxonomic unit (OTU) clustering and taxonomic analyses were performed using CLC Genomics Workbench V.10.1.1 and CLC Microbial Genomics Module V.2.5 (Qiagen). Sequences were first trimmed to remove 13 bases at the 5′ terminal position and merged considering the alignment scores as follows: mismatch cost of 2, gap cost of 2, zero maximum unaligned end mismatches and minimum score of 30.
After merging, sequences were clustered into OTUs at 97% sequence similarity level. The most abundant sequences were selected as representative of each cluster and then assigned to a taxonomy level using default values and the Greengenes Database 2013 release. Low depth samples (less than 2000 sequences per sample) were removed from the analysis. Alpha diversity indexes (Simpson, Shannon and total OTU number) were calculated. Bray-Curtis and unweighted UniFrac metric were used to calculate intersample diversity (beta diversity).
Samples were rarefied using QIIME 1.9, and multiple comparisons and statistical analyses were performed using CLC Genomics Workbench V.10.1.1 and CLC Microbial Genomics Module V.2.5 (Qiagen). A Negative Binomial GLM model was used to obtain maximum likelihood estimates for an OTU’s log-fold change between two conditions, and the Wald test was used to determine significance. False discovery rate (FDR) was performed to correct p values.
Data availability
The datasets generated during the current study are available in the National Center for Biotechnology Information (NCBI) BioProject Repository https://www.ncbi.nlm.nih.gov/bioproject under BioProject ID PRJNA596776.
Pathogen analysis using quantitative PCR
Levels of Campylobacter jejuni, C. concisus, C. lari, C. upsaliensis, C. coli, Escherichia coli enteroinvasive, Salmonella enterica, Shigella dysenteriae and Yersinia enterocolitica were determined in appendiceal DNAs from subjects with acute non-perforated appendicitis and control subjects by qPCR, using species-specific primers. The primers used for the analysis of Campylobacter species are described in table 1. qPCR for the remaining gastrointestinal (GI) pathogens was performed using a customised 96-well qPCR array provided by Qiagen (Custom Catalogue number CBAID00160), containing pre-prepared primers and probes designed to detect E. coli enteroinvasive (Cat. number BPVF00473A), S. enterica (Cat. number BPID00302A), S. dysenteriae (Cat. number BPID00307A) and Y. enterocolitica (Cat. number BPID00357A).
Primers used to quantify Campylobacter species in appendiceal tissues from subjects with appendicitis and control subjects
qPCR for all Campylobacter species was performed on a Bio-Rad CFR96 Real-Time System (Bio-Rad) as follows: 1.2 µL of 10 µM forward primer, 1.2 µL of 10 µM reverse primer, 10 µL of iQ SYBR Green Supermix (Bio-Rad), 5.6 µL of PCR grade water and 2 µL of template DNA, for 95°C for 10 min, 40 cycles of 95°C for 30 s, 58°C for 1 min, 72°C for 1 min, and a final step of 95°C for 10 s. qPCR for E. coli enteroinvasive, S. enterica, S. dysenteriae and Y. enterocolitica was performed according to the manufacturer’s instructions (Qiagen) on a QuantStudio 6 Flex System (Thermo Fisher Scientific, Waltham, Massachusetts, USA).
qPCR statistical analysis
Mann-Whitney tests and construction of Receiver Operator Characteristic (ROC) curves and graphs were performed using GraphPad Prism V.7.02 (GraphPad Software, La Jolla, California, USA).
Results
Study samples
Samples of inflamed tissues from 50 consecutive cases of acute non-perforated appendicitis in female subjects were included in this study (mean age=34 years, range 18–53 years). Samples of non-inflamed healthy appendiceal tissues were obtained from 50 gender-matched control subjects who underwent incidental appendectomy during gynaecological surgery. Of these 50 control samples, 8 were excluded due to the presence of IBS on chart review. Similarly, one sample was excluded due to IBD and six were excluded due to incomplete medical records. As a result, 35 control samples were included for microbiome analysis (mean age=32, range 18–47 years).
Diversity of bacterial species in phylum Proteobacteria is altered in acute appendicitis compared with healthy appendiceal tissues
A total of 50 samples from areas of inflamed tissue from subjects with acute non-perforated appendicitis and 35 samples from areas of non-inflamed tissue from control subjects with healthy appendiceal tissues were sequenced. Three control samples generated less than 2000 reads and were excluded from the analysis.
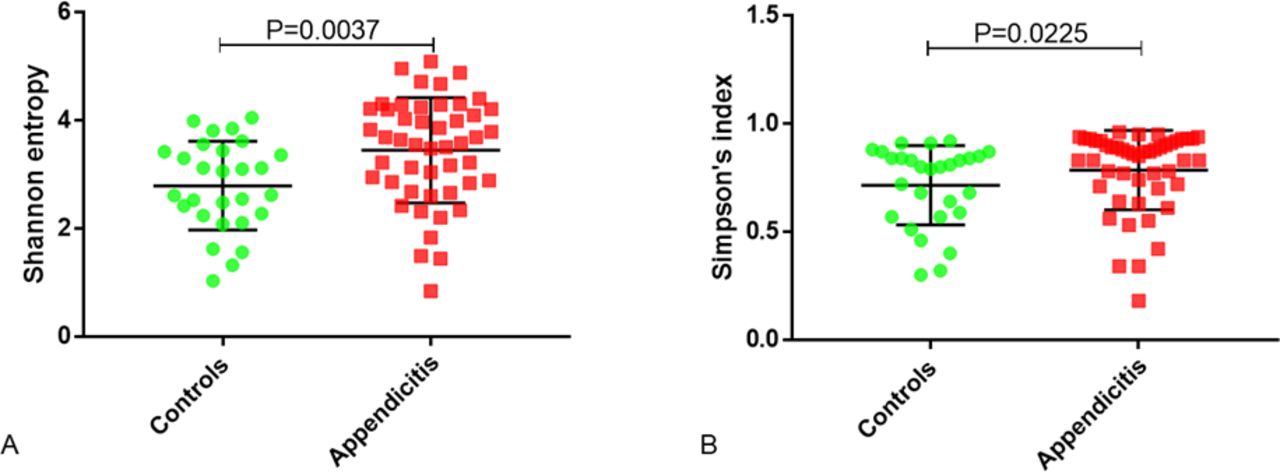
Alpha diversity was similar in the appendiceal microbiomes of appendicitis samples compared with healthy controls, with no significant differences between the two groups for Shannon entropy (p=0.2708), Simpson’s index (p=0.5219), or the total number of observed OTUs (p=0.3749) (figure 1). Alpha diversity indices were then calculated for each of the major phyla detected in each group. The results revealed that both the Shannon and Simpson’s diversity indices for phylum Proteobacteria were increased in the appendicitis group compared with controls (Shannon p=0.0037 and Simpsons p=0.0225) (figure 2).

Alpha diversity indices (A—Shannon entropy, B—Simpson’s index and C—total operational taxonomic unit (OTU) number) for the appendiceal microbiomes in inflamed tissues from acute appendicitis samples (n=50, red) and in non-inflamed tissues from healthy appendiceal control samples (n=32, green). P values were calculated using the Mann-Whitney test.

Alpha diversity indices for phylum Proteobacteria (A—Shannon entropy and B—Simpson’s index) in inflamed tissues from acute appendicitis samples (red) and in non-inflamed tissues from healthy appendiceal control samples (green). P values were calculated using the Mann-Whitney test.
The appendiceal microbiome in acute non-perforated appendicitis has a distinct microbial signature associated with the phylum Proteobacteria
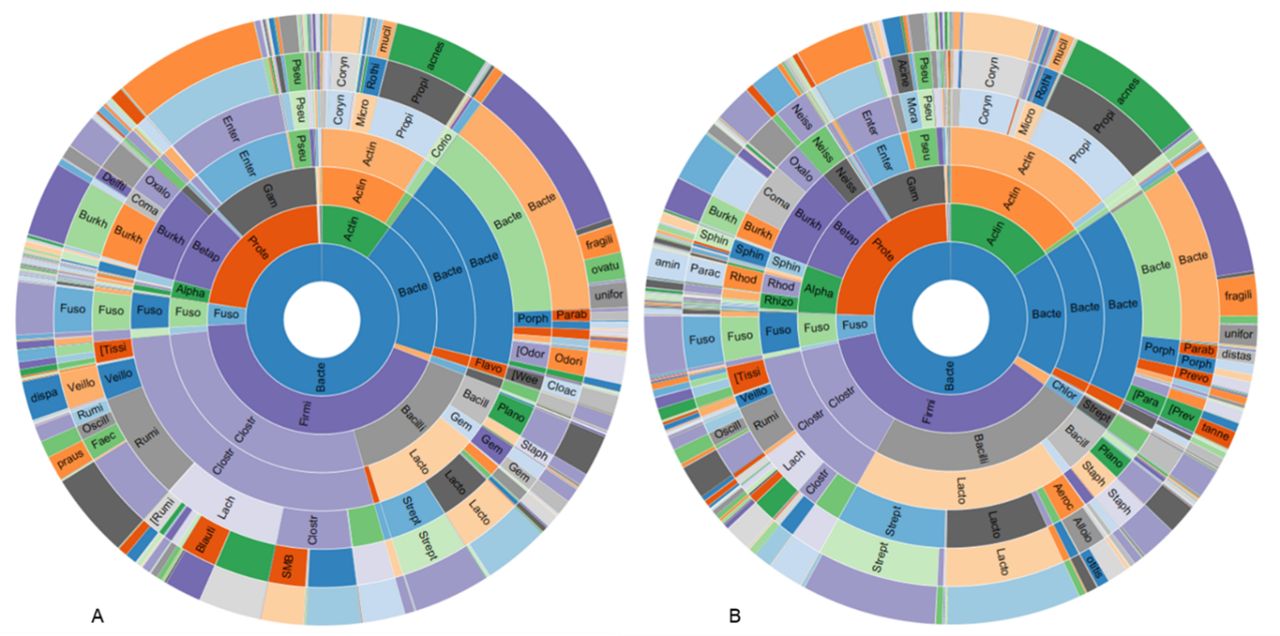
The RAs of the major phyla in healthy appendiceal control tissues were as follows: Firmicutes~43%, Proteobacteria~22%, Bacteroidetes~20%, and Actinobacteria 10%. The RAs of the major phyla in acute appendicitis tissues were similar, with Firmicutes~37%, Proteobacteria~24%, Bacteroidetes~18%, and Actinobacteria 16%, but there was a slight decrease in Firmicutes (Log2 fold change (FC)=−0.69, FDR p value=1.89E-3) and increase in Cyanobacteria (FC=0.88, FDR p value=0.029) in appendicitis samples compared with controls (figure 3).
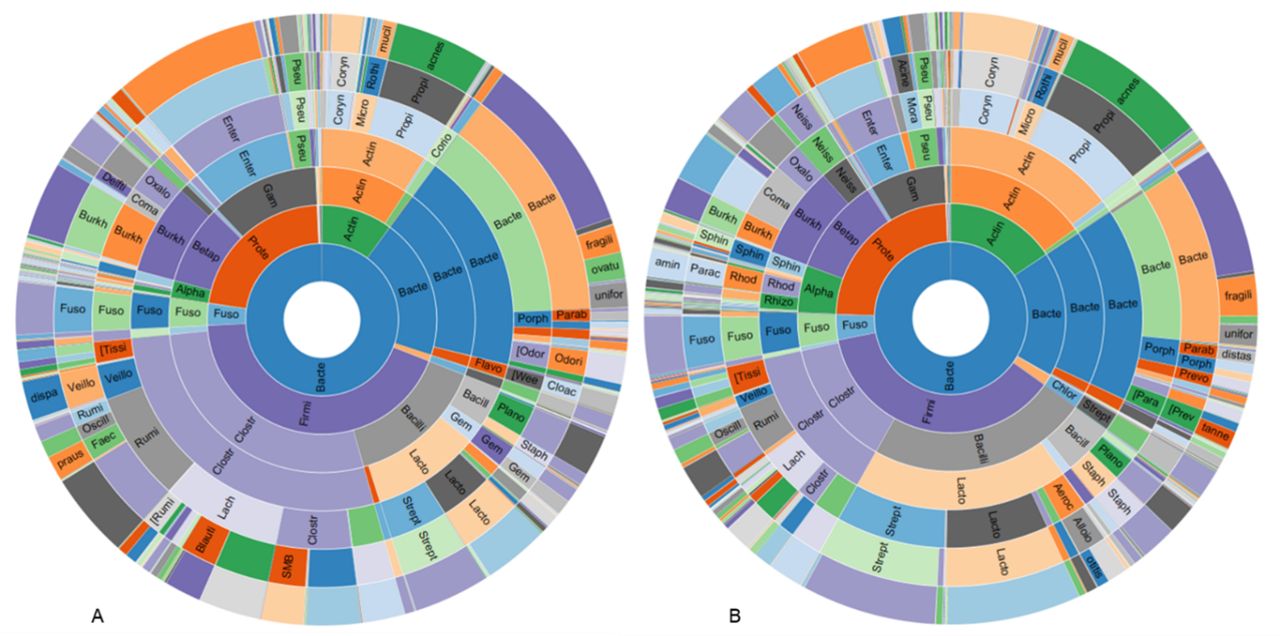
Sunburst representations of the appendiceal microbiome profiles in control subjects with healthy appendices (A) and in subjects with acute appendicitis (B).
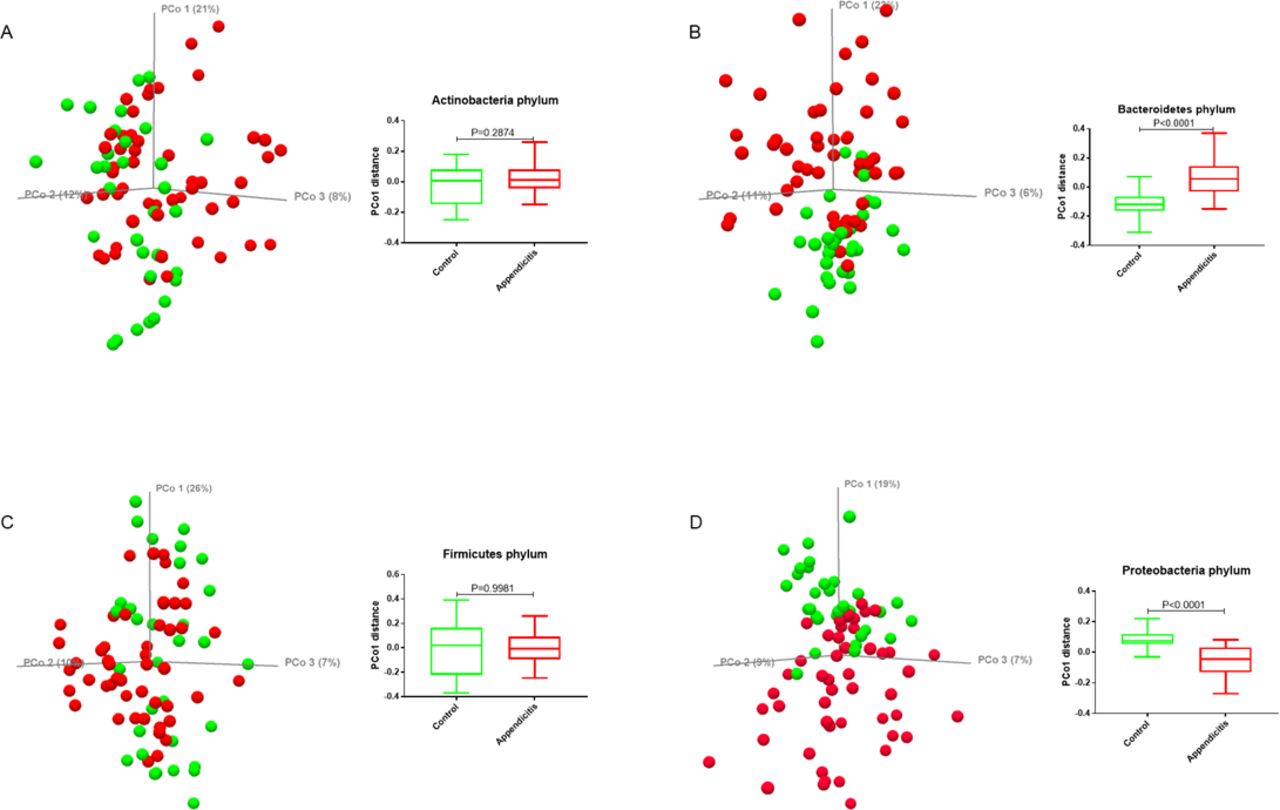
Principal coordinate analysis (PCoA) of the Bray-Curtis distance revealed a difference in beta diversity between the acute appendicitis and healthy appendiceal samples (PCo1<0.0001 and PCo2=0.0126) (figure 4). To define the major phyla associated with these differences in beta diversity, separate PCoA analyses were performed for each phylum (figure 5). The unweighted UniFrac distances of the microbial communities in the phyla Bacteroidetes and Proteobacteria were different in subjects with appendicitis versus healthy controls (p<0.0001) (figure 5B and D, respectively). In contrast, the UniFrac distances of the microbial communities in the phyla Actinobacteria and Firmicutes were not statistically different between the two groups (figure 5A and C, respectively).

(A) Principal coordinate analysis (PCoA) plot of the Bray-Curtis distance (beta diversity) between acute appendicitis (red) and healthy appendiceal samples (green). (B, C) Boxplots showing the statistical differences in PCo1 and PCo2 distances between the two groups. P values were calculated from the Mann-Whitney test.

Principal coordinate analysis (PCoA) plot and differences in the unweighted UniFrac distances between acute appendicitis (red) and healthy appendiceal (green) samples for each of the four major phyla. (A) Actinobacteria, (B) Bacteroidetes, (C) Firmicutes, and (D) Proteobacteria. P values were calculated from the Mann-Whitney test.
Alterations in phylum Proteobacteria in the appendix of subjects with appendicitis
Analyses at the class level revealed that there was an increased RA of the class Alphaproteobacteria (~21%, FC=1.31, FDR p value=0.03), and a smaller increase in the class Epsilonproteobacteria (~1%, FC=0.25, p value=0.029, FDR p value>0.05), in acute appendicitis samples compared with controls. The RAs of the classes Betaproteobacteria (~43%), Gammaproteobacteria (~34%), and Deltaproteobacteria (~1%) were not significantly different between the two groups (figure 6).

Sunburst representations of phylum Proteobacteria in appendiceal tissues from control subjects with healthy appendices (A) and from subjects with acute appendicitis (B).
At the family level, a total of 29 phylum Proteobacteria families were altered in acute appendicitis samples compared with controls (Mann-Whitney test p value<0.05 and Wald test FDR p value<0.05) (figure 6). Of these, 5 exhibited decreased RA in appendicitis samples, including Burkholderiaceae (FC=−1.81, FDR p value=0.01) and Enterobacteriaceae (FC=−1.34, p value=0.009, FDR p value>0.05), and 23 exhibited increased RA, including the families Moraxellaceae (FC=1.27, FDR p value=0.02), Pasteurellaceae (FC=1.77, FDR p value=0.02), Neisseriaceae (FC=1.73, FDR p value=0.01), Comamonadaceae (FC=1.96, FDR p value=7.89E-5), Sphingomonadaceae (FC=1.02, p value=0.012, FDR p value>0.05) and Campylobacteraceae (FC=0.60, p value=0.042, FDR p value>0.05) (figure 6).
Extending these analyses to the genus level revealed alterations in 16 genera from the phylum Proteobacteria in acute appendicitis samples compared with controls (figure 6). Of these, 2 exhibited decreased RA in acute appendicitis samples, including an unknown genus from the family Burkholderiaceae (FC=−2.37, p value=0.001) and an unknown genus from the family Enterobacteriaceae (FC=−0.76, p value=0.005), and 14 exhibited increased RA, including Neisseria (FC=2.35, p value<0.0001), Acinetobacter (FC=2.07, p value<0.0001) and Campylobacter (FC=1.58, p value=0.042).
Campylobacter jejuni is associated with acute non-perforated appendicitis
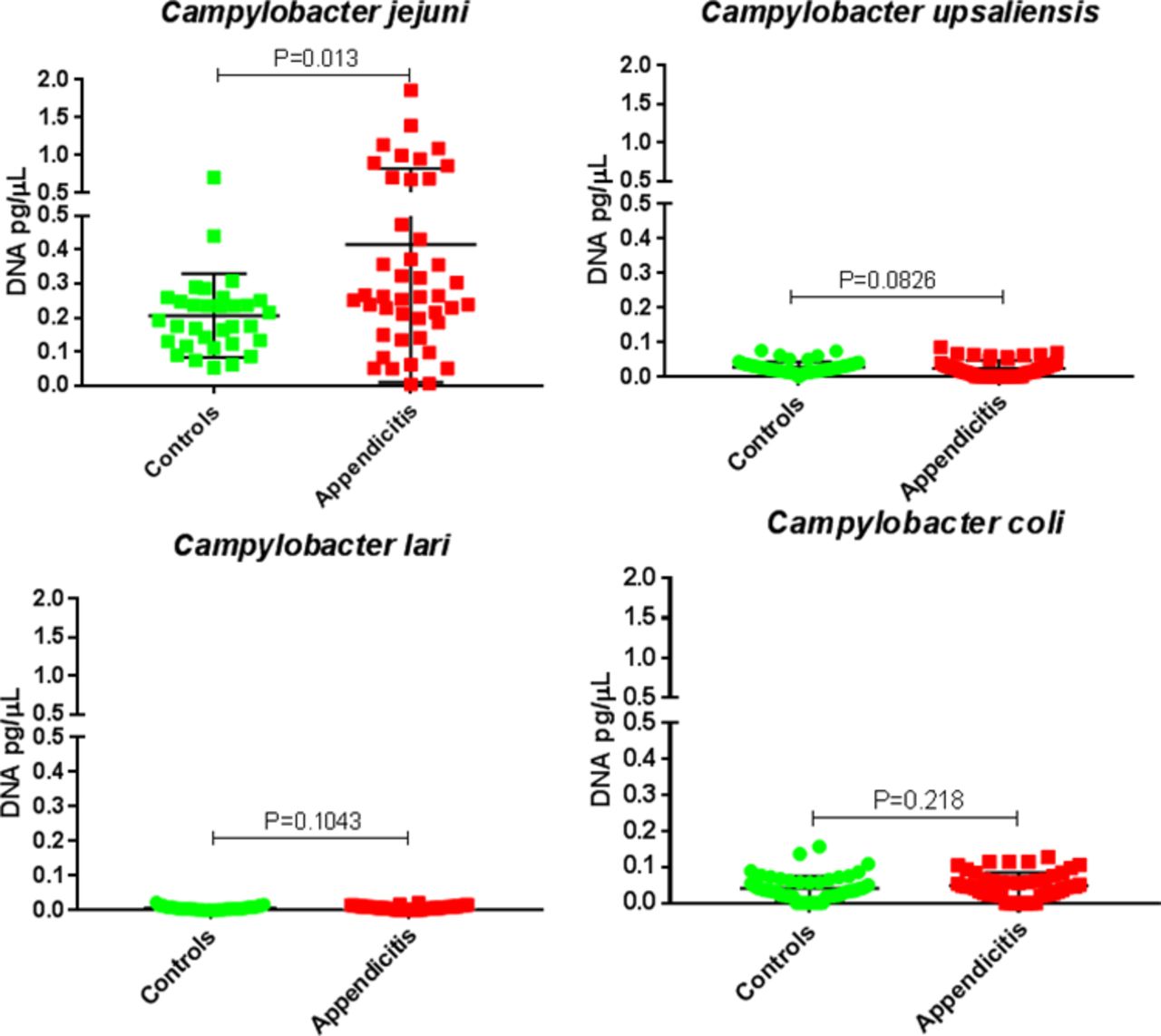
Given the increased RA identified in genus Campylobacter in appendicitis samples compared with controls, and previous reports suggesting potential links between Campylobacter species and appendicitis,20–22 the levels of five Campylobacter species (C. jejuni, C. upsaliensis, C. lari, C. coli and C. concisus) in appendiceal tissues were determined by qPCR, using the remaining DNA from samples from 44 subjects with acute appendicitis and 32 controls. No DNA from C. concisus was detected in appendiceal tissues from either group. Levels of C. jejuni DNA were significantly higher in tissues from subjects with appendicitis (median=0.26 pg/µL) than in controls (median=0.18 pg/µL) (p=0.013). There were no statistically significant differences between the two groups for the other Campylobacter species (figure 7).
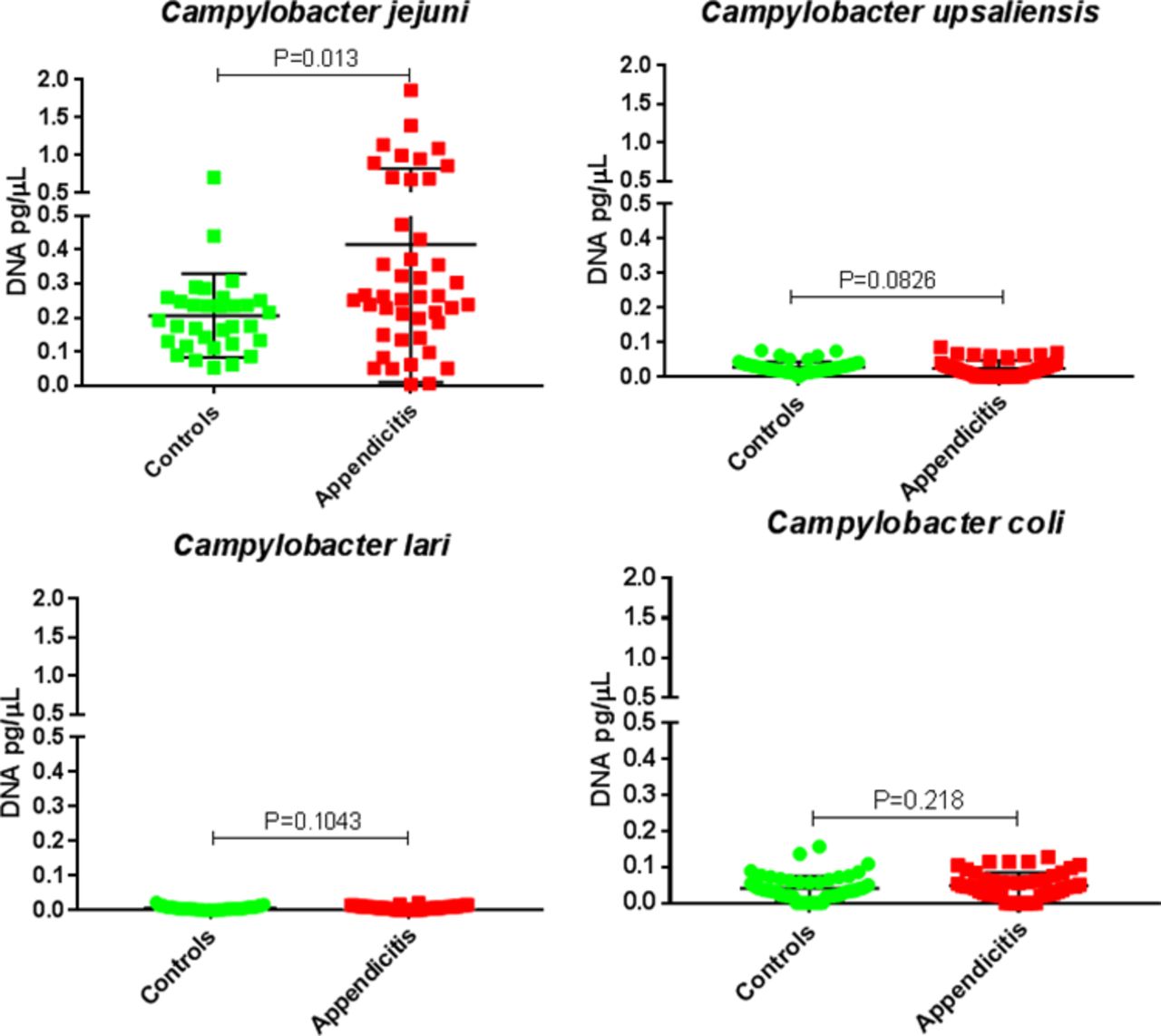
Levels of DNA from Campylobacter species in appendiceal tissues from controls (green) and subjects with appendicitis (red).
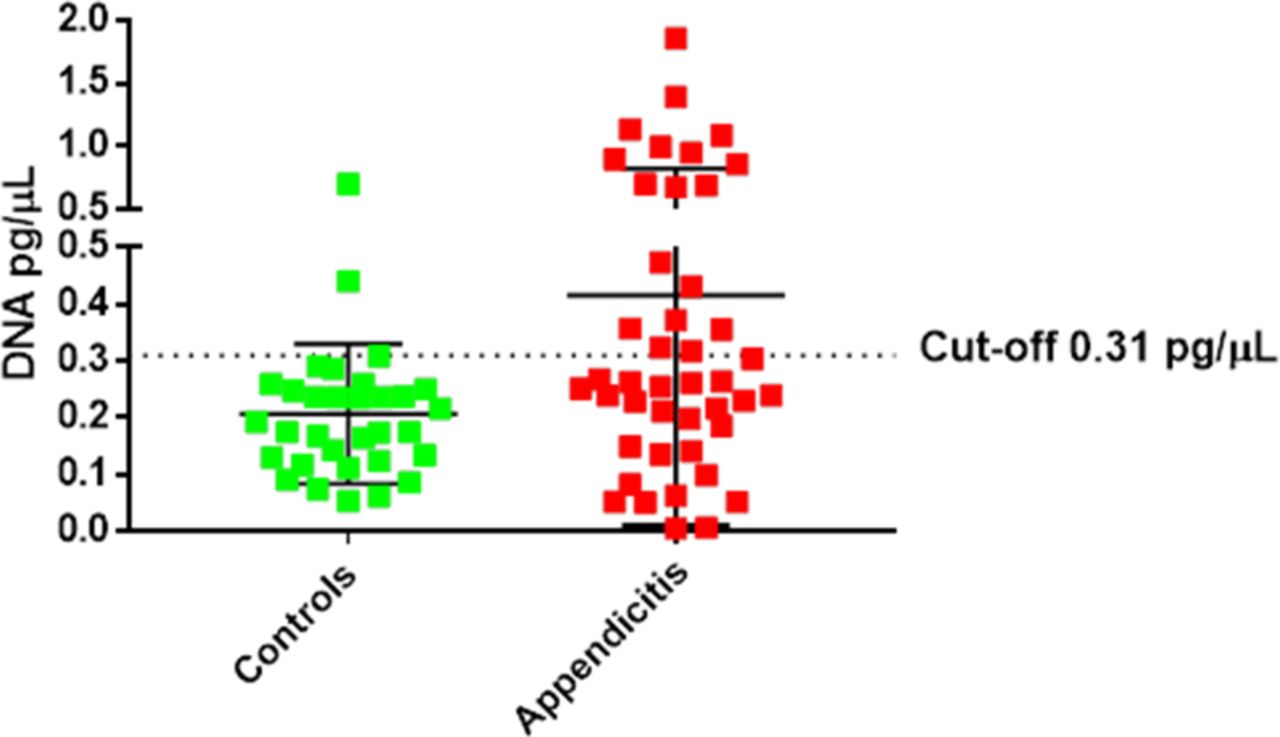
Further analyses revealed that there was an association between the levels of C. jejuni present in appendiceal tissues and acute appendicitis. Using a cut-off of 0.31 pg/µL of C. jejuni DNA, 40% of appendicitis cases and 6% of controls were positive for C. jejuni, giving a specificity of 93.7% (95% Cl 79.2 to 99.2), sensitivity of 40.9% (95% Cl 24.7 to 54.5) and OR of 10.38 (Fisher’s p value=0.0006, 95% Cl 2.3 to 47.4) (figure 8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Using a cut-off of 0.31 pg/µL Campylobacter jejuni DNA levels identified subjects with appendicitis with a specificity of 93.7%, sensitivity of 40.9% and OR of 10.38.
No significant levels of GI pathogens in appendiceal tissues from subjects with acute appendicitis versus controls
In addition to determining the levels of the Campylobacter species described above, the levels of four other common GI pathogens, E. coli enteroinvasive, S. enterica, S. dysenteriae and Y. enterocolitica, were also compared in appendiceal tissues from subjects with acute appendicitis and controls. No detectable levels of these pathogens were observed in appendiceal tissues from either group.
Discussion
This study compares the appendiceal microbiome in subjects who presented with acute non-perforated appendicitis versus control subjects with healthy appendiceal tissues, using an optimised technique which allows for the extraction of bacterial DNA from FFPE histological slides of sufficient quantity and quality for 16S rRNA gene sequencing. Specifically, we compared areas of inflamed appendiceal tissue from adult subjects with acute appendicitis to areas of non-inflamed appendiceal tissue from control subjects who underwent incidental appendectomy during gynaecological surgery. The results revealed significant differences in the appendiceal microbiome between these two groups, including a significant difference in overall beta diversity driven by differences in the phyla Bacteroidetes and Proteobacteria. There were also significant differences between the groups within phylum Proteobacteria. These included increased RA of the families Burkholderiaceae, Moraxellaceae and Campylobacteraceae, and of an unknown genus from family Burkholderiaceae as well as the genera Acinetobacter and Campylobacter, in inflamed tissues from acute appendicitis subjects compared with non-inflamed control tissues. qPCR subsequently revealed that levels of C. jejuni, but not other Campylobacter species tested, were elevated in appendiceal tissues from subjects with acute appendicitis when compared with controls with healthy appendices. This is consistent with earlier reports in the literature suggesting a role for C. jejuni in acute appendicitis.20–22 Moreover, we identified a threshold level for C. jejuni in appendiceal tissues that was associated with appendicitis.
Recently, there has been an interest in using antibiotics for the treatment of stable cases of acute appendicitis. Typical regimens include intravenous antibiotics for 1–3 days followed by oral antibiotics for up to 10 days, but the choice of antibiotics is not standardised.25 Our confirmation of C. jejuni as an organism of interest in appendicitis suggests that targeted treatment with antibiotics that eliminate C. jejuni, such as ciprofloxacin, could prove efficacious. Although larger studies would be required to confirm this, the use of antibiotic treatments could have significant healthcare and economic benefits. Appendectomy still is the most common abdominal surgical emergency in the world, and while the incidence of appendicitis has for the most part stabilised in industrialised Western countries, it continues to rise in newly industrialised countries in Asia, South America and Africa.3 Impacting the rate of appendectomies through the use of targeted antibiotic therapies could therefore result in considerable healthcare savings worldwide.
Our identification of C. jejuni as a potential pathogen in acute appendicitis could also be useful in assessing patients prior to surgery. Our analyses reveal that using a cut-off of 0.31 pg/µL of C. jejuni DNA resulted in the identification of acute appendicitis with a specificity of 93.7%, sensitivity of 40.9% and an OR of 10.38. If elevated C. jejuni levels are also found in stool samples, this could be used in the future to determine the presence of C. jejuni in uncomplicated suspected appendicitis prior to surgery. Metabolomic analyses might also facilitate the identification of a serum biomarker which could be used in rapid bedside diagnostic testing. Again, further prospective studies will be needed to explore these possibilities.
In addition to C. jejuni, associations between other bacteria and appendicitis have also been described. Early culture-dependent studies suggested a role for Bacteroides fragilis in acute appendicitis with septic complications,18 and bacteria that are normally found in the GI tract, such as E. coli and Streptococcus spp, have also been recovered from appendiceal tissues.19 More recent studies have attempted to characterise the appendiceal microbiome using next-generation sequencing. These include a small study of seven patients that found Firmicutes, Proteobacteria and Bacteroidetes to be the most abundant phyla in the appendiceal microbiome, which is consistent with our findings, and which also noted a high prevalence of the genus Fusobacterium in appendiceal samples.5 Although that study lacked a control group, making the identification of causative organisms difficult, the high prevalence of Fusobacterium is consistent with results from other studies. Zhong et al found an increased abundance of Fusobacterium spp and reduced abundance of Bacteroides spp in paediatric appendicitis versus non-appendicitis cases,7 and also found increased abundances of oral bacteria such as Porphyromonas, Parvimonas and Gemella in the appendicitis cases. Another paediatric study found increases in 12 bacterial taxa in appendicitis subjects versus those with normal appendices, including Peptostreptococcus, Bilophila, Bulledia, Fusobacterium, Parvimonas and Porphyromonas.6 Schulin et al performed both culture and 16S rRNA sequencing of appendiceal samples, again from paediatric subjects, and identified 11 genera by culture, of which Escherichia showed the highest abundance, and 109 genera by 16S sequencing, of which Fusobacterium showed the highest abundance, with Porphyromonas and Parvimonas also identified.8 We did not find significant differences in the levels of any of these taxa in appendicitis versus control tissues in our study, which may be due to the fact that we used adult subjects, or that we specifically compared inflamed appendicitis tissues to non-inflamed controls.
This study has some limitations. First, our study was limited to female subjects, as the control samples were obtained from females undergoing gynaecological surgeries with incidental appendectomy. While the gender of the host would likely have little effect on the microbiota of the appendix, we cannot discount the possibility of gender being a confounding variable in the results of this study. It should also be noted that, per institutional policy, preoperative antibiotics are given to all subjects immediately before surgery, including both appendectomies and gynaecological surgeries. While we do not anticipate that this treatment could have had an immediate effect on the appendiceal microbiome, again we cannot rule out confounding effects.
In summary, we have shown that in over a third of cases of acute appendicitis, there is a significant increase in the levels of the pathogen C. jejuni. None of the other Campylobacter species or other common GI pathogens tested were found to be increased in appendicitis or control cases. The identification of a pathogen such as C. jejuni as common in appendicitis may help to guide therapeutic options in the acute setting. This, in turn, could have significant implications in treating and reducing the most common cause of emergent abdominal surgery in the USA and globally.
References
Footnotes
Contributors MP, AR and RM conceived and designed the study. SJO, AR, NP and JZ identified potential subjects and matched controls and procured samples, and JZ identified relevant tissue areas for analysis. TS-E and GMB wrote the supporting regulatory documents, and TS-E assisted with database searches. MP, GL, SC, MJV-M, IL, BC, GP, WM, SW and GMB performed the experiments, and GGSL, AR and MP analysed the data. GGSL, MP, GMB, RM, and MP wrote, reviewed and edited the manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The study protocol was approved by the Institutional Review Board at Cedars-Sinai Medical Center.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available in a public, open access repository. The datasets generated during the current study are available in the National Center for Biotechnology Information (NCBI) BioProject Repository https://www.ncbi.nlm.nih.gov/bioproject under BioProject ID PRJNA596776.