Article Text

Abstract
Objective We aimed at the identification of genetic alterations that may functionally substitute for CTNNB1 mutation in ß-catenin-activated hepatocellular adenomas (HCAs) and hepatocellular carcinoma (HCC).
Design Large cohorts of HCA (n=185) and HCC (n=468) were classified using immunohistochemistry. The mutational status of the CTNNB1 gene was determined in ß-catenin-activated HCA (b-HCA) and HCC with at least moderate nuclear CTNNB1 accumulation. Ultra-deep sequencing was used to characterise CTNNB1wild-type and ß-catenin-activated HCA and HCC. Expression profiling of HCA subtypes was performed.
Results A roof plate-specific spondin 2 (RSPO2) gene rearrangement resulting from a 46.4 kb microdeletion on chromosome 8q23.1 was detected as a new morphomolecular driver of β-catenin-activated HCA. RSPO2 fusion positive HCA displayed upregulation of RSPO2 protein, nuclear accumulation of β-catenin and transcriptional activation of β-catenin-target genes indicating activation of Wingless-Type MMTV Integration Site Family (WNT) signalling. Architectural and cytological atypia as well as interstitial invasion indicated malignant transformation in one of the RSPO2 rearranged b-HCAs. The RSPO2 gene rearrangement was also observed in three β-catenin-activated HCCs developing in context of chronic liver disease. Mutations of the human telomerase reverse transcriptase promoter—known to drive malignant transformation of CTNNB1-mutated HCA—seem to be dispensable for RSPO2 rearranged HCA and HCC.
Conclusion The RSPO2 gene rearrangement leads to oncogenic activation of the WNT signalling pathway in HCA and HCC, represents an alternative mechanism for the development of b-HCA and may drive malignant transformation without additional TERT promoter mutation.
- molecular genetics
- hepatocellular carcinoma
- adenoma
- molecular carcinogenesis
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Hepatocellular adenoma (HCA) represents a benign liver tumour that can be divided into molecular subtypes with variable risk of malignant transformation into hepatocellular carcinoma (HCC). Mutations of the CTNNB1 gene currently characterise the group of ß-catenin-activated HCAs (b-HCA), which carry an increased risk of malignant transformation, a process in which TERT promoter mutations were associated with the last step of the adenoma-carcinoma transition. CTNNB1 mutations also belong to the most prevalent genomic alterations in human HCC.
What are the new findings?
A roof plate-specific spondin 2 (RSPO2) gene rearrangement resulting from a 46.4 kb microdeletion on chromosome 8q23.1 leads to upregulation of RSPO2 protein and activation of WNT signalling, as indicated by diffuse nuclear accumulation of CTNNB1 and upregulation of WNT target genes. This constellation was demonstrated for cases of b-HCA, b-HCA with transition to HCC and HCC in the context of chronic liver disease. Malignant transformation does not require an additional TERT promoter mutation, which is associated with malignant transformation of b-HCAs with CTNNB1 mutation, but coexistence of RSPO2 rearranged and CTNNB1-/TERT promoter-comutated HCC clones may occur as a consequence of tumour heterogeneity. Thus, the RSPO2 rearrangement is an oncogenic driver functionally substituting for activating CTNNB1 mutations in both b-HCA and HCC. The frequency of RSPO2-rearrangement was 1.6% (3/185) in HCA, 18% (3/17) in b-HCA and 0.6% (3/474) in HCC, respectively.
Significance of this study
How might it impact on clinical practice in the foreseeable future?
A preclinical mouse model of colon cancer showed that intestinal RSPO2-fusion tumours are entirely WNT dependent and can be treated by inhibition of WNT secretion (eg, porcupine inhibitor). Thus, such an approach might also be promising in RSPO2 rearranged liver tumours. We provide a promising molecular target and the respective biomarker to select HCC patients that may benefit from WNT secretion inhibition.
Introduction
Hepatocellular neoplasia in adults can be divided in hepatocellular adenoma (HCA) and hepatocellular carcinoma (HCC). While HCA comprise a group of benign liver tumours that mainly develop in females without pre-existing liver disease in the context of oral contraception,1 HCC represents one of the most common cancers worldwide, has a male predilection, occurs in the setting of chronic liver diseases (eg, chronic viral hepatitis and steatohepatitis) and has a poor prognosis.2 3
Activating mutations of the CTNNB1 gene are among the most frequent somatic alterations in human HCC4 5 and additionally have been described to characterise a subgroup of HCA, which carry an increased risk of malignant transformation into HCC (so-called ß-catenin-activated HCA [b-HCA]).1 6–9 About half of all b-HCA reveal additional features of inflammatory HCA (b-IHCA).1 10 Overall, the frequency of CTNNB1 mutation is higher in HCC (up to 35%) than in HCA (10%–15%).1 11 A recent large cohort study demonstrated that the type of CTNNB1 mutation determines the degree of ß-catenin activation in HCA and HCC. Most exon 3 mutations result in high activity, while mutations in exons 7 (K335) and 8 (N387), and S45 mutants in exon 3 lead to weaker pathway activation, as indicated by the degree of transcriptional activation of classical ß-catenin target genes like GLUL (coding for glutamine synthetase [GS]) and the leucine-rich repeat-containing G-protein-coupled receptor 5 (LGR5).7 The immunohistological GS expression pattern is considered a reliable marker for detection of b-HCA.12 13 Diffuse and strong GS expression was observed in liver tumours with high-activity CTNNB1 mutations, while the low-activity mutations (S45, K335 and N387) resulted in a weaker and more heterogeneous GS positivity. Interestingly, when the weak S45 mutant alleles were detected in HCC, they were duplicated in most cases leading to increased ß-catenin activity,7 suggesting that a critical level of (nuclear) ß-catenin activation is required for malignant transformation of hepatocytes. However, we noticed several cases of ß-catenin-activated HCA and HCC without detectable mutations in the CTNNB1 gene.
The roof plate-specific spondin (RSPO) protein family consists of four homologues (RSPO1, RSPO2, RSPO3 and RSPO4) that are evolutionarily conserved in vertebrates and involved in a broad range of developmental and physiological processes.14 RSPO ligands potentiate WNT/β-catenin signalling through binding to leucine-rich repeat-containing G protein-coupled receptors (LGR). This leads to increased WNT signalling as the transmembrane E3 ubiquitin ligases zinc and ring finger 3 and its homologue ring finger 43 (RNF43),15 16 which negatively regulate WNT/β-catenin signalling by promoting clearance of WNT receptors from the cell surface, are themselves displaced following RSPO binding to LGRs.17 18
Here, we describe a new molecular driver of β-catenin-activated HCA and HCC generated by a 46.4 kb microdeletion on chromosome 8q23.1 leading to a roof plate-specific spondin 2 (RSPO2) gene rearrangement that results in powerful activation of β-catenin signalling.
Material and methods
Human tissue samples
Between 2008 and 2018, 185 HCA from 173 patients were diagnosed and morphomolecularly subtyped in the Institute of Pathology, University Hospital Heidelberg. The male-to-female ratio was 1 to 15. The median age was 38 years (range 10–72). The specimens included 99 biopsies and 74 resections; 115 cases were received for second opinion. Liver adenomatosis was present in 14 patients, glycogenosis type 1 was known in three patients and familial adenomatous polyposis was noted in one patient. Tumour rupture occurred in four and use of anabolic steroids was known in two patients. Cases were reviewed by two expert hepatopathologists (TL and PS), and all HCAs were classified according to the current WHO classification adopting recent amendments.1 19–21 RSPO2 protein expression was analysed in normal adult liver, HCA and well-differentiated HCC using a tissue microarray (TMA; online supplementary table 1). For statistical comparison of HCA subgroups, the Wilcoxon rank-sum test was used (IBM SPSS Statistics V.24 software). P<0.05 was considered significant.
Supplementary file 1
Features of RSPO-rearranged liver tumours
Since January 2018, molecular profiling of HCC patients eligible for further systemic treatment after failure of sorafenib was performed at our institution. This cohort included six male cases without detectable CTNNB1 mutation. The median age of these patients was 60 years (range 26–75 years). Two cases each occurred in the setting of chronic hepatitis B and chronic alcohol abuse, respectively. One case was associated with non-alcoholic steatohepatitis, and in one case, the aetiology was unknown. Liver cirrhosis was present in four out of six cases. To evaluate the frequency of β-catenin-activated HCC with a wild-type CTNNB1 gene, a second TMA containing human HCC (n=468) was used. The main characteristics of this cohort are summarised in online supplementary table 2.
Immunohistochemistry and digital pathology
Immunohistochemistry was performed on 3 µm sections using an automated immunostainer (Ventana BenchMark Ultra, Roche Diagnostics, Basel, Switzerland) as described in the online supplementary material, which also contains the method used for quantitation of β-catenin-positive nuclei.
Sanger sequencing of the CTNNB1 gene (exons 3, 7 and 8)
One hundred ng DNA were used for PCR amplification, and all PCR reactions were run at a final volume of 25 µL. Exons 3, 7 and 8 of CTNNB1 were evaluated for the presence of mutations by PCR amplification and direct sequencing. The primer sequences are shown in online supplementary table 3
DNA extraction, library preparation and semiconductor sequencing
Next-generation sequencing (NGS) is a high-throughput technology that allows for the rapid and simultaneous evaluation of many genes facilitated by sequencing of millions of short nucleic acid sequences in parallel. Ion semiconductor sequencing is an NGS method detecting hydrogen ions that are released during the building of a complementary strand based on the sequence of a template strand (so-called sequencing-by-synthesis). DNA and RNA extraction, library preparation and semiconductor sequencing were conducted as described in theonline supplementary material.22 23
Anchored multiplex PCR (AMP)-based NGS translocation detection
AMP-based translocation detection is an NGS technology, which uses unidirectional gene-specific primers to facilitate open-ended amplification and thus the detection of novel rearrangements in RNA derived from formalin-fixed paraffin-embedded (FFPE) samples in combination with molecular barcodes to enable unique molecule counting and error correction. For enriched cDNA library preparation, the multiplex PCR-based Ion Torrent AmpliSeqTM technology (Thermo Fisher Scientific) with the Archer FusionPlex Solid Tumor assay (ArcherDX, Boulder, Colorado, USA) was used as described in the online supplementary material.
Expression profiling using digital probe-based analysis
Expression profiling of 14 HCA cases (6 b-HCA, 3 b-IHCA, 3 inflammatory HCA [IHCA] and 2 HNF1-α- inactivated HCA [H-HCA]) including one RSPO2-rearranged b-HCA and RSPO2-rearranged b-IHCA, as well as one RSPO2-rearranged HCC and three normal liver tissues was performed using the nCounter platform with the nCounter PanCancer Pathways Panel (online supplementary NanoString Technologies, Seattle, Washington, USA) according to the manufacturer’s instruction. Briefly, 200 ng of total FFPE RNA was hybridised to the nCounter probe sets for 16 hours at 65°C. Samples were processed using an automated nCounter Sample Prep Station (online supplementary NanoString Technologies). Cartridges were subsequently imaged on an nCounter Digital Analyzer (online supplementary NanoString Technologies) set at 490 fields of view. Reporter counts were collected using NanoString’s NSolver software V.3.0. Raw counts are shown as online supplementary material. For comparison, all samples were normalised against the positive control and five housekeeping genes (TLK2, ERCC3, PIAS1, DNAJC14 and SLC4A1AP) selected according to the manufacturer’s instructions. For all further pathway analysis, the fold change was determined by normalisation against the mean expression level of the gene of interest in three normal liver tissues. Pathway analysis were carried out using Ingenuity Pathway Analysis (release October 2018; Qiagen, Hilden, Germany).
Supplementary file 2
Reanalysis of TCGA HCC data
Gene expression data and mutational calls of 373 HCCs of the LIHC TCGA cohort were downloaded from the Cancer Genome Data Server (CGDS; http://www.cbioportal.org) using the CGDS R package. The cut-off point of 2.57 to differentiate CTNNB1 wild-type HCC with high respectively low RSPO2 expression was determined by fitting a mixture model of two normal distributions using Cutoff Finder.24 Violin plots of gene expression data were generated using the ggplot2 R package.25 Significance of gene expression changes was assessed with Welch’s t-test.
Results
Frequency, clinical and molecular features of β-catenin-activated HCA subtypes
Between 2009 and 2018, 185 HCA from 173 patients were histologically diagnosed and subtyped at our institution (figure 1). These included 53 H-HCA (29%), 97 IHCA (52%), three b-IHCA (2%), 14 b-HCA (8%) and 18 unclassified HCA (UHCA, 10%). The median age of patients with b-HCA was 28 years (range 10–45). These included nine women and two men. Use of anabolic steroids was known in two male patients. Information on tumour size was available from eight patients (73%). The mean size of b-HCA was 8.1 cm (range 6–13 cm). Six of 13 cases (46%) were diagnosed by liver biopsy. Concerning b-IHCA, the median age of patients was 41 years (range 25 to 57; one male, two females). The mean size of b-IHCA was 5.7 cm (range 3.2–10 cm). 1/3 cases (33%) was diagnosed by liver biopsy. H-HCA (median 43 years, range 19–62, p=0.001) and IHCA (median 38 years, range 21–72, p=0.004) patients were significantly older than b-HCA patients. In addition, H-HCA patients were significantly older than UHCA patients (median 31, range 21–46, p=0.008).

Frequency of HCA subtypes and molecular features of b-HCA and b-IHCA. APC, adenomatosis polyposis coli; b-IHCA, ß-catenin-activated inflammatory HCA; b-HCA, ß-catenin-activated HCA; H-HCA, HNF1-α- inactivated HCA; HCA, hepatocellular adenoma; IHCA, inflammatory HCA; RSPO2, r oof plate-specific spondin 2; UHCA, unclassified HCA.
The mutation spectrum of b-HCA was as follows: mutations in exon 3 of the CTNNB1 gene were present in 10 cases (71%), including two cases harbouring an S45 mutation. In addition, two b-HCA cases revealed an S335 mutation (exon 7). No CTNNB1 mutation was detected in two b-HCAs. All b-IHCA patients were CTNNB1 wild type, but one patient was known to suffer from familial adenomatous polyposis due to an APC germline mutation. The activation modes of WNT/β-catenin signalling are summarised in online supplementary table 4. Malignant transformation was seen in three b-HCA harbouring a CTNNB1 exon 3 mutation and one b-IHCA with a wild-type CTNNB1 gene.
Molecular changes in β-catenin-activated HCA without conventional CTNNB1 mutation
By immunohistochemical detection of diffuse and strong GS expression, we detected two b-HCAs and three b-IHCAs in which no mutation was found in exons 3, 7 and 8 of the CTNNB1 gene. One b-IHCA harboured a germline APC mutation in a patient with known FAP. The remaining four adenomas were submitted to combined targeted DNA and RNA sequencing using the Oncomine Comprehensive Assay V.3. While no mutations were observed in CTNNB1, RNF43 or other driver genes, a recurrent 46.4 kb microdeletion on chromosome 8q23.1 leading to the connection of a short interspersed nuclear element (SINE) to exon 2 of RSPO2 was detected in two b-HCAs and one b-IHCA (figure 2A or B). Due to insufficient tumour tissue available for molecular diagnostics, one b-IHCA could not be analysed by RNA sequencing. Thus, the mode of activated WNT/β-catenin signalling remained elusive in this case. The RSPO2 rearrangement was independently validated by AMP-based NGS translocation detection (figure 2A). Overall, the frequency of RSPO2 rearrangement was 18% (3/17) in all variants of β-catenin-activated adenomas. All RSPO2-rearranged cases showed some cytological atypia and strong homogeneous GS expression. In contrast to β-catenin-activated HCA resulting from a mutation in exons 3, 7 and 8 of the CTNNB1 gene, which usually show nuclear β-catenin expression only in a minority of tumour cells, nuclear β-catenin accumulation was present in most tumour cells of RSPO2-rearranged HCA (figure 3 or 4, online supplementary figure 1). Quantitation of β-catenin positive nuclei in β-catenin-activated HCA revealed a significantly higher percentage of β-catenin positive nuclei in RSPO2-rearranged adenomas compared with HCA carrying a CTNNB1 mutation in exon 3 (mean 56.5%±21.9 vs 6.0%±4.0, p=0.02; online supplementary table 4 and figure 2).
Supplementary file 3

(A) The connection of exon 2 of RSPO2 to the 5′-upstream located short interspersed nuclear element (SINE) was demonstrated by semiconductor sequencing using the Oncomine Comprehensive Assay version 3 (OCAv3) and AMP-based NGS translocation detection using Archer FusionPlex Solid Tumor assay. (B) Graphical illustration of the genomic and the transcript structure resulting from the detected 46.4 kb microdeletion at chromosome 8q23.1. (C) Diffuse upregulation of RSPO2 protein in an RSPO2 rearranged HCA and (D) very faint periportal expression in the corresponding non-neoplastic liver tissue (200-fold). AMP NGS, anchored multiplex PCR-based next-generation sequencing translocation detection; RefSeq, Reference Sequence Genome Reference Consortium Human Build 37 (GRCh37); RSPO2, roof plate-specific spondin.

Expression pattern of CTNNB1 (upper row) and GS (lower row) in b-HCA differ between different modes of WNT signalling activation. b-HCA, ß-catenin-activated hepatocellular adenoma; GS, glutamine synthetase.

(A) Highly differentiated hepatocellular tumour with architectural atypia as indicated by pseudogland formation and focally more than two cells wide trabeculae (original magnification 100-fold). (B) No upregulation of SAA expression (100-fold). (C) Diffuse and strong GS positivity (100-fold) and (D) prominent nuclear β-catenin accumulation (400-fold). Overall, the picture is typical of b-HCA. b-HCA, ß-catenin-activated hepatocellular adenoma; GS, glutamine synthetase; SAA, serum amyloid A.
In addition, a fusion of exon 1 of the oestrogen receptor 1 and exon 8 of coiled-coil domain containing 170 gene (ESR1(E2)-CCDC170(C8)) was detected in one case. Comparing all RSPO2-rearranged b-HCA, this tumour showed prominent fatty change and steatohepatitic features, which were not seen in the other three cases. The clinicopathological parameter and additional molecular alterations detected by sequencing are detailed in table 1.
RSPO2 protein expression in HCA subtypes and normal liver
The translational start site of RSPO2 is located in exon 2; thus, this rearrangement is predicted to allow for the expression of a full-length RSPO2 protein. Consequently, we observed upregulation of RSPO2 protein in all HCA with a rearranged RSPO2 gene (figure 2C), while RSPO2 staining was restricted to hepatocytes surrounding portal tracts in the corresponding normal liver tissue (figure 2D).
To characterise the RSPO2 expression in normal adult liver tissue and highly differentiated hepatocellular neoplasm in more detail, a TMA containing samples from liver resections was used. Overall, diffuse RSPO2 expression was detected in 3% of normal adult liver tissues (n=5/191), while RSPO2 expression was restricted to periportal hepatocytes in 90% of cases (n=171/191; online supplementary figure 3A). In the remaining cases no RSPO2 staining was seen in normal liver tissue. In HCA, no staining was seen in 72% (n=41/57), focal RSPO2 positivity was seen in 12% (n=7/57; including one IHCA, one b-IHCA, two H-HCA and 3 UHCA) and a diffuse expression was seen in 16% (n=9/57) of HCA. These included all RSPO2-rearranged HCA (n=2) and all HCA with CTNNB1 exon 3 mutation (n=4; online supplementary figure 3C). In addition, each one H-HCA, IHCA and UHCA revealed a diffuse RSPO2 protein staining. In well-differentiated HCC, no RSPO2 protein was detected in 62% (n=16/26), focal staining was seen in 31% (n=8/26) and diffuse RSPO2 expression was seen in 8% of HCC (n=2/26).
Supplementary file 5
Malignant transformation of RSPO2 gene rearranged liver tumours
Malignant transformation into HCC was observed in 1/53 H-HCA (2%), 1/97 IHCA (1%), 1/3 b-IHCA (33%) and 3/14 b-HCA (21%), respectively. Malignant transformation occurred more frequently in b-HCA compared with other subtypes (b-HCA vs IHCA, p=0.006; b-HCA vs H-HCA, p=0.027; H-HCA vs IHCA, p>0.05).
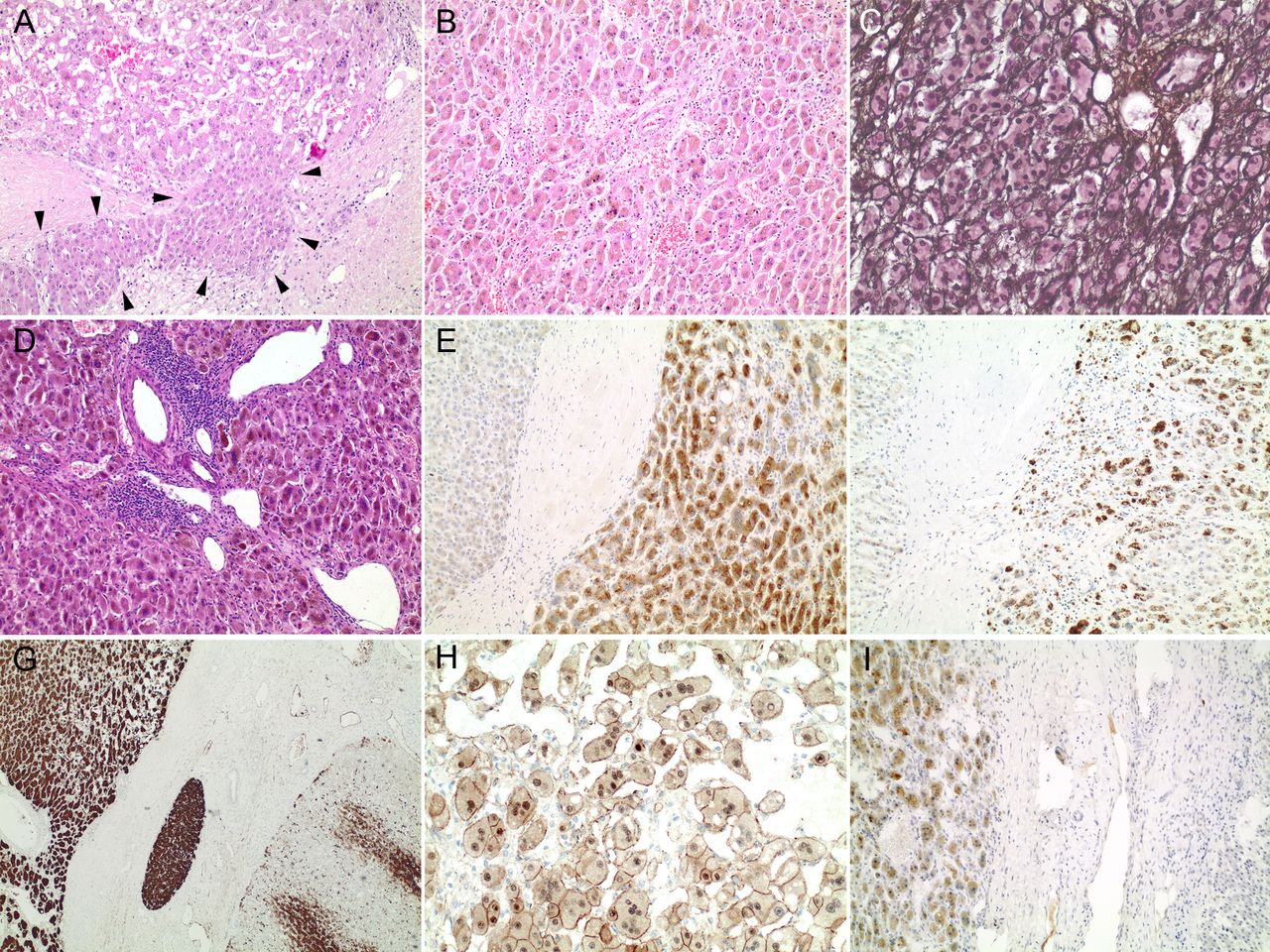
As the strong nuclear β-catenin accumulation in RSPO2-rearranged HCA indicated a high β-catenin activation status, we reasoned that these adenomas may be prone to malignant transformation. Indeed, one of the RSPO2 positive b-IHCA showed focal pseudocapsule formation with evidence of interstitial invasion (figure 5), upregulation of GPC3 protein and a diffuse capillarisation of the intralesional sinusoids (CD34) indicating malignant transformation to HCC. No TERT promoter mutation was detected in this tumour (and any of the other CTNNB1 mutation negative HCA cases). As CTNNB1 mutations belong to the most frequent somatic mutations in HCC as well,4 5 we reanalysed HCC cases that underwent molecular profiling to evaluate a personalised targeted approach after failure of sorafenib treatment at our institution. In one of six CTNNB1-wild-type HCCs present in this collective, an identical RSPO2 rearrangement was detected. Here, tumour development occurred in a 26-year-old male patient in the setting of liver cirrhosis due to chronic hepatitis B (figure 6). The clinicopathological data of this HCC cohort are summarised in online supplementary table 5. To estimate the frequency of RSPO2 rearranged HCC, another large cohort of human HCC (n=468; online supplementary table 2) was evaluated for nuclear CTNNB1 expression, which was detected in 8% (36/468) of tumours. In 3% of HCC (12/468) an at least moderate nuclear accumulation of CTNNB1 (IRS≥6) was recorded, and these cases were submitted to targeted sequencing. A CTNNB1 mutation (exon 3) was detected in 9/12 cases (75%), while an RSPO2 rearrangement was found in two samples (online supplementary table 6). Quantitation of β-catenin positive nuclei in β-catenin-activated HCA revealed a significantly higher percentage of β-catenin positive nuclei in RSPO2-rearranged HCC compared CTNNB1 exon 3 mutated HCC (mean 91.2%±6.8 vs 54.3%±10.1, p=0.03; online supplementary table 6 and figure 2).

(A) Highly differentiated hepatocellular tumour with focal interstitial invasion (arrow heads). Note: the higher degree of nuclear atypia of the invasive tumour cells (original magnification 100-fold). (B) Note isolated arteries, discrete microvesicular steatosis and areas of abrupt anaplasia (100-fold). (C) Focal loss of the reticulin network as well as nuclear atypia are evident (modified Gomori, 200-fold). (D) Typical histological features of inflammatory HCA like portal tract like structures with dilated vessels and aggregates of inflammatory cells are evident (100-fold). (E) CRP and (F) SAA are upregulated compared with the surrounding non-neoplastic liver tissue (left part; each 100-fold). (G) Diffuse and strong GS staining in tumour cells (upper left), while the surrounding liver tissue (lower right) reveals an orthotopic GS expression (40-fold). (H) Nuclear β-catenin accumulation (200-fold). (I) In addition, the tumour cells overexpress glypican-3 (100-fold). This histological picture is indicative of b-IHCA with malignant transformation in HCC. b-IHCA, ß-catenin-activated inflammatory HCA; GS, glutamine synthetase; HCC, hepatocellular carcinoma.

(A) Moderately differentiated HCC with trabecular growth pattern (original magnification 200-fold). (B) Chronic hepatitis B is evident by cytoplasmic retention of HBs antigen (200-fold). (C) Diffuse GS expression (200-fold) and D) both cytoplasmic and nuclear CTNNB1 accumulation (400-fold). HBs, hepatitis B surface antigen; HCC, hepatocellular carcinoma.
Interestingly, in one case, a CTNNB1 mutation and a TERT promoter mutation occurred together with an RSPO2 rearrangement. Noteworthy, the allele frequencies of the CTNNB1 and the TERT mutation were approximately 17%, while all other mutations and polymorphism showed an allele frequency >40% (eg, ATM 46%). In one case, neither a CTNNB1 mutation nor an RSPO2 rearrangement, but mutations of the cbl proto-oncogene and neurofibromin 1 genes were detected, both of which may activate CTNNB1 signalling. In the last patient, the quality of the extracted nucleic acids from FFPE tissue did not allow for a successful genetic analysis.
RSPO2 rearrangement leads to activation of CTNNB1-target genes
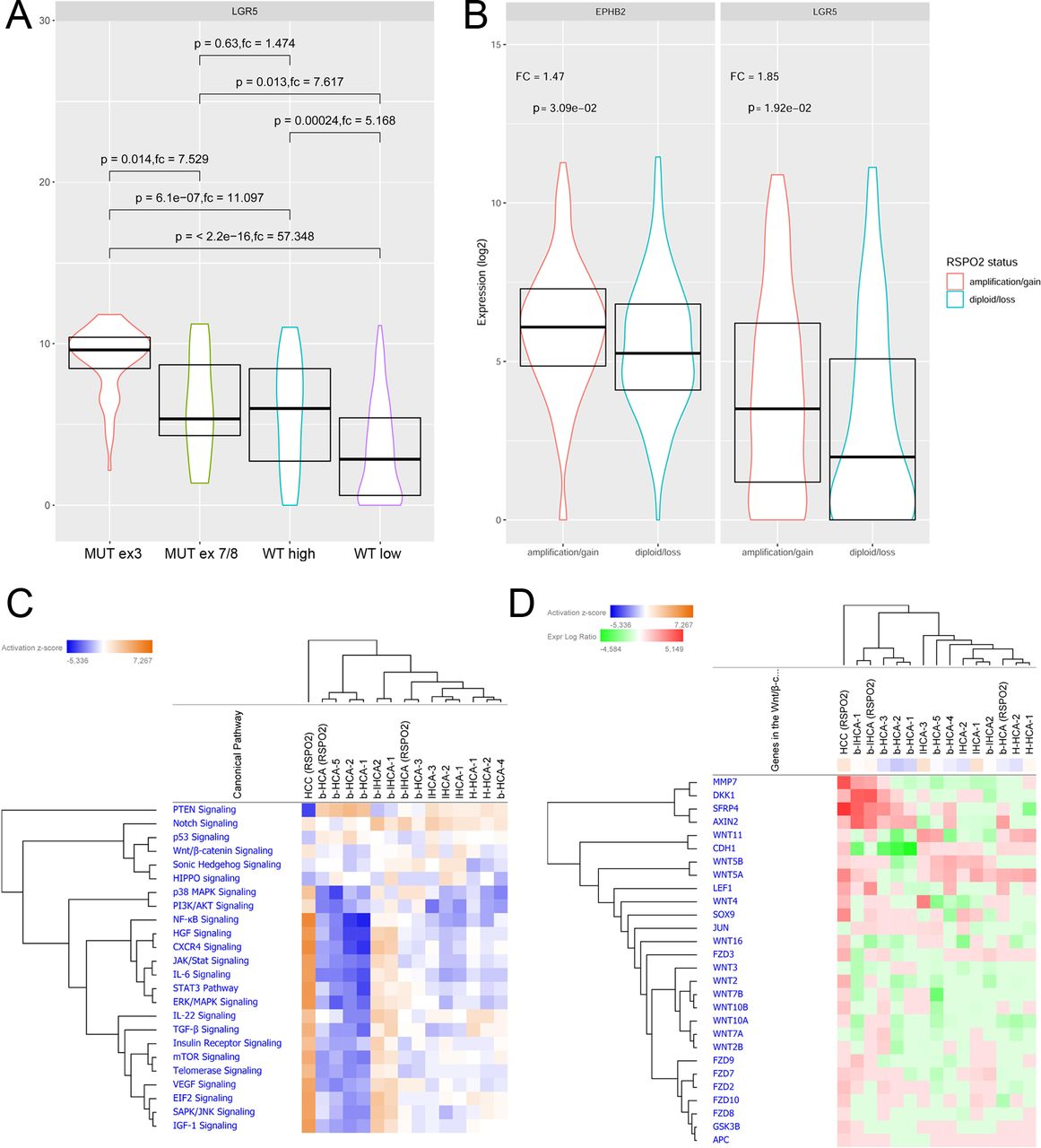
The relevance of RSPO2 upregulation for the pathogenesis of human HCC was further supported by reanalyses of the TCGA HCC cohort.5 As expected, high mRNA expression of WNT target genes including GLUL (8.1-fold change, p<0.001), LGR5 (31-fold change, p<0.001) and TBX3 (6.1-fold change, p<0.001) was strongly associated with β-catenin mutated HCCs. Of note, the RSPO2 expression was significantly higher in HCCs with CTNNB1 mutation in exon three compared with exon 7/8 mutated cases (GLUL: 10.2-fold change, P<0.001; LGR5: 7.5-fold change, P=0.01; TBX3: 2.9, P=0.003). Additionally, CTNNB1 wild-type HCCs with high RSPO2 expression showed significantly higher expression of β-catenin target genes than RSPO2 low expressing HCCs without CTNNB1 mutation (GLUL: 2.2-fold change, p=0.001; LGR5: 5.2-fold change, p<0.001; TBX3: 1.8, p=0.02; figure 7A, online supplementary figure 4). Further analysis revealed an RSPO2 gene amplification in 16% of HCCs of the TCGA cohort. CTNNB1 wild-type HCCs with copy number gains at the RSPO2 gene locus revealed significantly increased activation of β-catenin target genes (p>0.05; figure 7B).
Supplementary file 6

{kind=link}
![[gutjnl-2018-317632supp003.jpg]](https://gut.bmj.com/content/gutjnl/68/7/1287/DC3/embed/inline-supplementary-material-3.jpg?download=true){kind=link}
{kind=link}
{kind=link}
{kind=link}
![[gutjnl-2018-317632supp004.jpg]](https://gut.bmj.com/content/gutjnl/68/7/1287/DC4/embed/inline-supplementary-material-4.jpg?download=true){kind=link}
{kind=link}
{kind=link}
![[gutjnl-2018-317632supp005.jpg]](https://gut.bmj.com/content/gutjnl/68/7/1287/DC5/embed/inline-supplementary-material-5.jpg?download=true){kind=link}
{kind=link}
(A) Expression of the CTNNB1-target gene LGR5 in human HCCs (TGCA cohort). LGR5 expression is higher in HCC with CTNNB1 expression in exon 3 compared with exon 7/8. In addition, CTNNB1-wild-type HCC with high RSPO2 mRNA levels reveal higher LGR5 expression compared with RSPO2 low expressing HCC. (B) An increased copy number of the RSPO2 gene is associated with upregulation of β-catenin target genes in CTNNB1-wild-type HCC. (C) Unsupervised hierarchical cluster analysis of liver cancer-associated pathways in HCA subtypes and one RSPO2-rearranged HCC. (D) Heatmap of genes belonging to the WNT/β-catenin pathway in these tumours. fc, fold change; HCA, hepatocellular adenoma; HCC, hepatocellular carcinoma; MUT ex3, CTNNB1 mutation in exon 3; MUT ex7/8, CTNNB1 mutation in exon 7/8; RSPO2, roof plate-specific spondin; WT high, CTNNB1 WT, high RSPO2 expression; WT low, CTNNB1 WT, low RSPO2 expression.
To get more insights into the biology of RSPO2-rearranged HCA and HCC, the expression of 770 genes from 13 cancer-associated pathways was determined using the online supplementary NanoString Technologies. As shown in figure 7C, the RSPO2-rearranged hepatocellular adenomas clustered together the respective CTNNB1-mutated adenoma subtypes. We did not detect a significant difference in expression of genes belonging to the WNT/β-catenin pathway between RSPO2-rearranged and CTNNB1-mutated HCA (p>0.05; figure 7D; online supplementary figure 5).
Supplementary file 7
![[gutjnl-2018-317632supp006.jpg]](https://gut.bmj.com/content/gutjnl/68/7/1287/DC6/embed/inline-supplementary-material-6.jpg?download=true){kind=link}
Discussion
The WNT/β-catenin pathway is a major regulator of liver development, zonation and regeneration.26–29 Under physiological conditions, WNT signalling is active in hepatocytes surrounding central veins in adult liver, while RSPO2 expression is restricted to periportal hepatocytes. As RSPO2-amplified β-catenin signalling requires the presence of WNT ligands and LRP6 expression, which are restricted to pericentral hepatocytes in healthy liver,14 30 it seems likely that RSPO2 does not contribute to hepatic WNT signalling under quiescent conditions.
Activated WNT signalling has been reported in up to 15% of HCA, which can be further subdivided into b-HCA or b-IHCA. In our series, activated WNT signalling was found in 10% of HCA (8% b-HCA and 2% b-IHCA, respectively). Although individual cases have been reported in the context of familial adenomatous polyposis coli (due to germ-line mutations of the adenomatous polyposis coli gene)31 as present in one patient with b-IHCA in this series, CTNNB1 mutations are considered the driving force of the vast majority of β-catenin-activated HCAs.1Compared with other HCA subtypes, b-HCAs have an increased risk for malignant transformation,1 6–9 which was confirmed in our series and emphasises that the reliable identification of b-HCA and b-IHCA subtypes among all HCAs has particular clinical relevance.
Chromosome rearrangements are a common oncogenic driving event in human tumours. DNAJB1-PRKACA translocations are the molecular driver in the fibrolamellar subtype of HCC,32 33 and INHBE-GLI1 rearrangements have been recently reported in a small subgroup of HCA with activated sonic hedgehog signalling,1 thus chromosomal rearrangements may also drive the development of certain HCA subtypes. Here, we identified a recurrent RSPO2 gene rearrangement due to a 46.4 kb microdeletion on chromosome 8q23.1 as the most prevalent cause of activated β-catenin signalling in HCA with wild-type CTNNB1 gene as demonstrated by targeted ultra-deep sequencing and independent validation using AMP-based NGS translocation detection. An identical microdeletion was previously described in colorectal cancer.34 Here, it was demonstrated that upstream of RSPO2, a 351 bp SINE was fused to exon 1 of the eukaryotic translation initiation factor 3 subunit E (EIF3E) gene. The high degree of DNA and RNA fragmentation in our formalin-fixed paraffin-embedded samples did not allow obtaining an SINE-spanning EIF3E-RSPO2 amplicon, and the GC-rich Alu sequences of the SINE hampered the primer design downstream of EIF3E. Thus, we were not able to prove the presence of the EIF3E gene upstream of the SINE sequence. EIF3E-RSPO2 gene rearrangements resulting from small deletions on chromosome 8q23.1 were predicted to produce a functional RSPO2 protein driven by the EIF3E promoter,35 and upregulation of RSPO2 protein has been reported in EIF3E-RSPO2 positive tumours.36 While EIF3E-RSPO2 alterations occurred mutually exclusive with other alterations of the WNT/β-catenin pathway in colorectal cancer, we observed one HCC with co-occurence of RSPO2 rearrangement, CTNNB1 and TERT promoter mutation. Interestingly, the frequencies of the variant CTNNB1 and TERT alleles was identical but much lower compared with the read counts of the coexisting ATM mutation and all detected polymorphisms suggesting the coexistence of CTNNB1 and TERT promoter mutation-positive and RSPO2-rearranged tumour cell clones. Interestingly, EIF3E upregulation has been observed during liver regeneration,37 indicating that the EIF3E promoter is active in replicating hepatocytes. We observed an upregulation of the RSPO2 protein, diffuse nuclear accumulation of β-catenin and strong upregulation of the β-catenin-target gene GS in RSPO2-rearranged b-HCA supporting the possibility of an EIF3E-RSPO2 gene fusion (E1ins351R2) in these tumours as well.
RSPO2 gene rearrangements were associated with cytological and architectural atypia in all RSPO2-rearranged HCA, and transition to HCC was observed in one b-HCA. Thus, HCA with RSPO2 gene rearrangements should be considered a variant of b-HCA requiring surgical resection due to an increased risk of malignant transformation.
Like CTNNB1-mutated HCA, RSPO2-rearranged HCA occurred mainly in young women. Moreover, we observed RSPO2-rearranged HCC that had developed in the setting of chronic liver disease, a situation in which malignant transformation of b-HCA is very unlikely indicating that RSPO2 rearrangements—similar to CTNNB1 mutations1 4–9—may drive both b-HCA and HCC development. Expectedly, the expression profile of the RSPO2-rearranged HCC was different from RSPO2-rearranged HCA (figure 7C). The oncogenic potential of RSPO2 upregulation was further supported by reanalysis of the TCGA data,5 which showed that high RSPO2 levels are associated with activation of β-catenin signalling. As RSPOs were shown to be WNT target genes in Xenopus,38 RSPO2 upregulation may provide a positive feedback to enhance WNT signalling. In line with this, strong activators of WNT signalling (eg, CTNNB1 exon 3 mutations) promoted higher RSPO2 levels than weaker inducers (eg, CTNNB1 exon 7 mutations). In addition, amplification of the RSPO2 gene locus, which has been reported in 3%–22% of human HCCs,39 40 was validated as an additional mechanism leading to activation of β-catenin signalling in the TCGA cohort emphasising the relevance of RSPO2 upregulation during hepatocarcinogenesis.
Although we observed upregulation of RSPO2 protein in all RSPO2-rearranged HCA, diffuse RSPO2 expression was also observed in most b-HCA with CTNNB1 exon 3 mutation. Thus, RSPO2 immunohistochemistry cannot be used as a reliable biomarker for the identification of RSPO2-rearranged HCA. In contrast, the quantitation of β-catenin positive nuclei in β-catenin-activated HCA and HCC corroborated that a prominent nuclear accumulation of CTNNB1 is a diagnostic hallmark of RSPO2-rearranged hepatocellular tumours. Thus, a prominent nuclear β-catenin accumulation (immunoreactive score (IRS) ≥6) should trigger further molecular analysis in case of HCA and HCC with a CTNNB1 wild-type gene (supplementary table 4 or 6 and figure 2).
It has been postulated that the transition from β-catenin-mutated HCA to HCC requires further oncogenic events (eg, TERT promoter mutation).8 Interestingly, our data suggest that a TERT promoter mutation is dispensable for malignant transformation of RSPO2-rearranged HCA, suggesting that the RSPO2 rearrangement might be a powerful oncogenic driver able to functionally substitute for both CTNNB1 and TERT promoter mutations.
Interestingly, the EIF3E–RSPO2 fusion was reported to be sufficient to initiate WNT-dependent tumour development in a CRISPR-based murine colon cancer model, and the WNT secretion inhibitor LGK974 could drive rapid tumour clearance in this model.41 Thus, inhibition of WNT secretion (eg, using a porcupine inhibitor) might be a promising target for the treatment of hepatic RSPO2-activated tumours, and our molecular diagnostic approach might serve as companion diagnostic to select HCC patients that may benefit from such a precision medicine concept. Considering the potential clinical impact of RSPO2 rearrangement in HCC, independent validation of our findings in large cohorts is desirable.
Supplementary file 4
![[gutjnl-2018-317632supp007.jpg]](https://gut.bmj.com/content/gutjnl/68/7/1287/DC7/embed/inline-supplementary-material-7.jpg?download=true){kind=link}
Acknowledgments
We would like to thank all our colleagues who have sent HCA cases for second opinion and without whose continuous support the present study would not have been possible. We are grateful to Lidia Jost, Diana Stürmer, Tina Uhrig, Kathrin Ridinger and Christiane Zgorzelski for excellent technical assistance. This work was supported by the tissue bank of the National Center for Tumor Diseases Heidelberg.
References
Footnotes
PS and AS contributed equally.
Contributors TL: experimental design, data interpretation, study supervision, drafting and critical revision of the manuscript; VE and RP: experimental design, data acquisition, data analysis and interpretation and revision of the manuscript; ON and MK: data acquisition, data analysis and revision of the manuscript; ER, SU and ZA: data analysis and interpretation and revision of the manuscript; MK, KHW, KB, AM, TFW, LW, BKS, AR and FS: data acquisition and revision of the manuscript; BB and SF: critical revision of the manuscript; JB: experimental design, data analysis and interpretation and drafting of the manuscript, PS and AS: experimental design, data analysis and interpretation, study supervision and critical revision of the manuscript.
Funding This work was supported by the Deutsche Forschungsgemeinschaft (LO 1676/2-1, LO 1676/4-1, SFB/TR209 project B08, to TL) and the German Cancer Consortium (DKTK, to PS and AS).
Competing interests None declared.
Ethics approval The retrospective study was approved by the local ethics committee of the University Hospital Heidelberg (S-346/2018).
Provenance and peer review Not commissioned; externally peer reviewed.
Patient consent for publication Not required.
Linked Articles
- Commentary